什么是 miRNA?
1993 年,科学家 Victor Ambros 等人在研究线虫时发现了一种名为
LIN-4 的基因,该基因表达一种小 RNA,可以抑制一种核蛋白
LIN-14 基因的表达[1]。这一小 RNA 就是第一个被报道的 miRNA (见往期推文:
)。此后,成千上万的 miRNA 在包括人类、小鼠、大鼠、斑马鱼、果蝇、水稻、拟南芥等几乎所有类群中陆续 “浮出水面”……
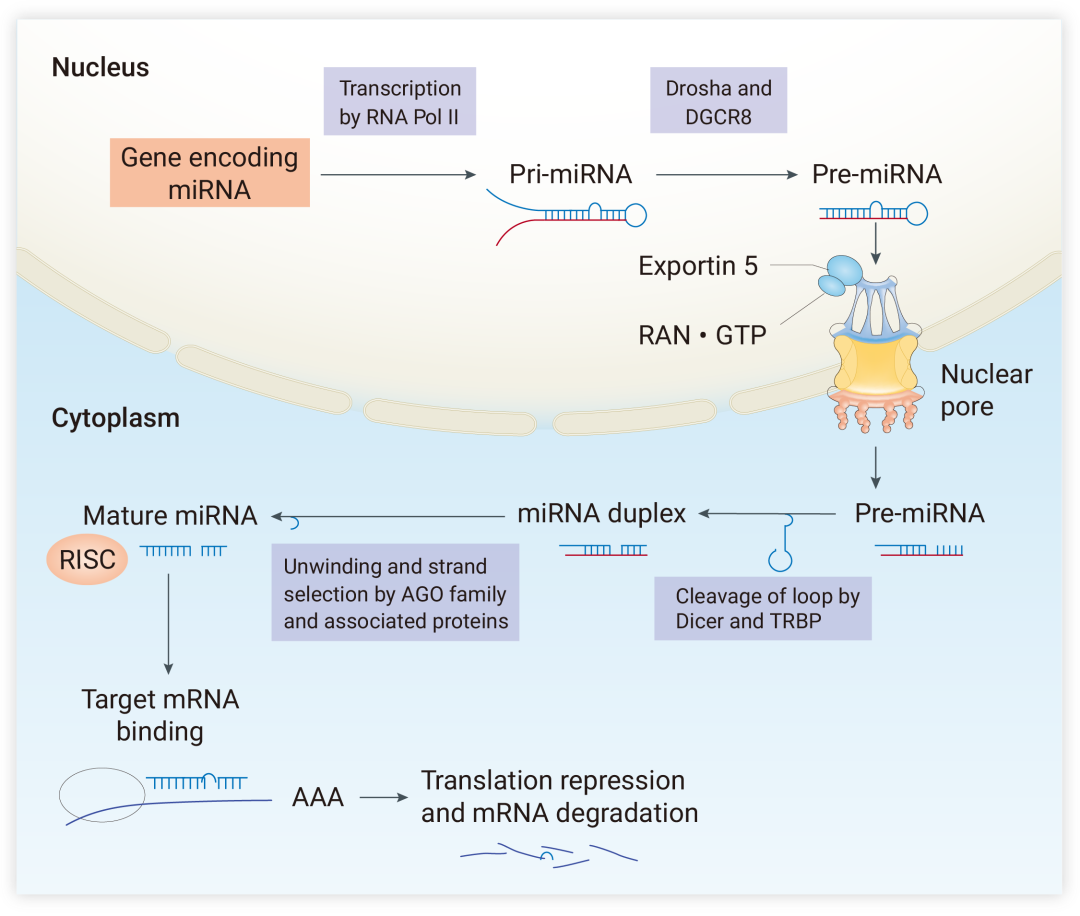
miRNA 是一种内源性的微小非编码 RNA,长度一般在 21-23 个核苷酸之间,具有在翻译水平调控基因表达的功能[2]。miRNA 的发生 “成长” 需要几个步骤:最初,miRNA 基因由 RNA 聚合酶 Ⅱ 转录,生成较长的前体 pri-miRNA,而后形成发夹状的 pre-miRNA[3,4]。最后在细胞质中被切割形成一个成熟的 miRNA 双链。至此,miRNA 从 >1000 个 nt 的 “大可爱” 摇身一变成为了仅约 22 个 nt 的 “小精灵”。成熟的 miRNA 会被装载到 Argonaute (AGO) 蛋白上,结合形成有活性的 RNA-诱导沉默复合物 (RNA-inducing silencing complex, RISC) 复合体,发挥其多样的调控功能 (图 1)[3]。
图 1. miRNA 的生物发生[3]
miRNA基因由RNA聚合酶Ⅱ 转录,生成较长的前体 pri-miRNA,然后与辅助因子 DGCR8 一起被核糖核酸酶 Drosha 切割,从而产生一个发夹状的 pre-miRNA。随后,在 Exportin 5-RNA•GTP 复合体介导下,pre-miRNA 从细胞核转移至细胞质中,并在细胞质中被核酸酶 Dicer 切割掉末端的茎环结构,形成一个成熟的 miRNA 双链。成熟的 miRNA 被装载到Argonaute(AGO) 蛋白上,结合形成 miRNA-诱导沉默复合物 (miRNA-inducing silencing complex,miRISC),AGO 蛋白会对 miRNA 两条链进行选择,5’端热稳定性较低的 miRNA 引导链 (miRNA guide strand) 优先被保留,另一条乘客链 (miRNA pass) 则被降解。
miRNA 与疾病调控
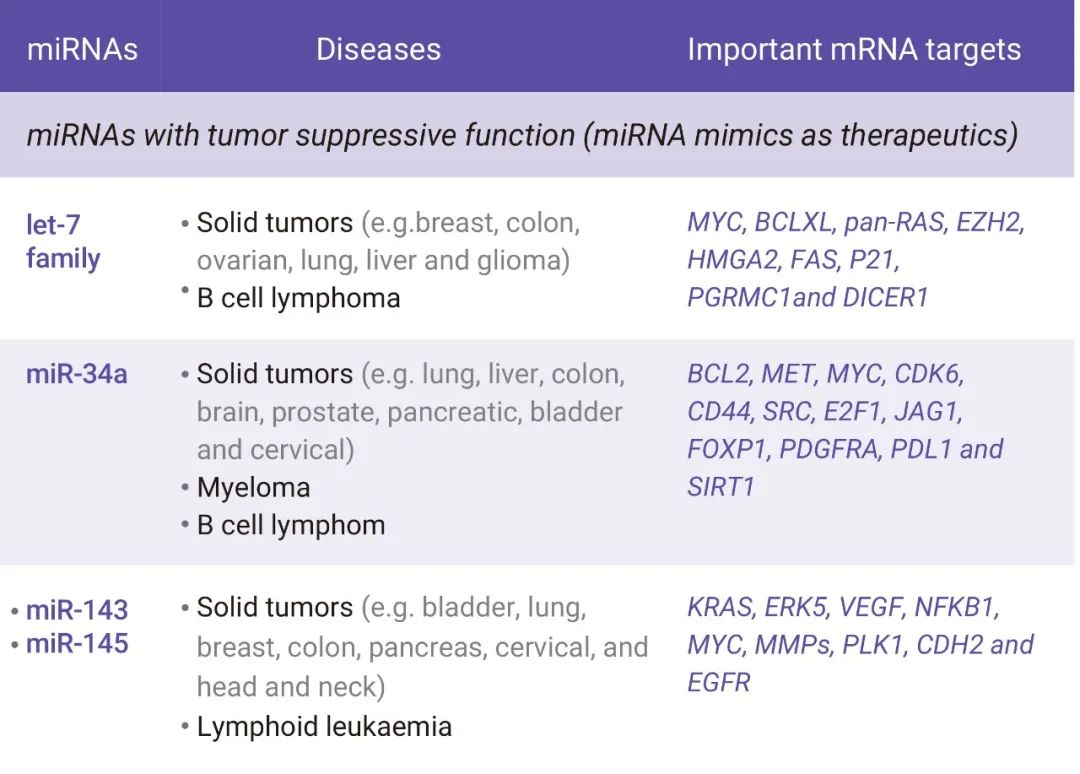
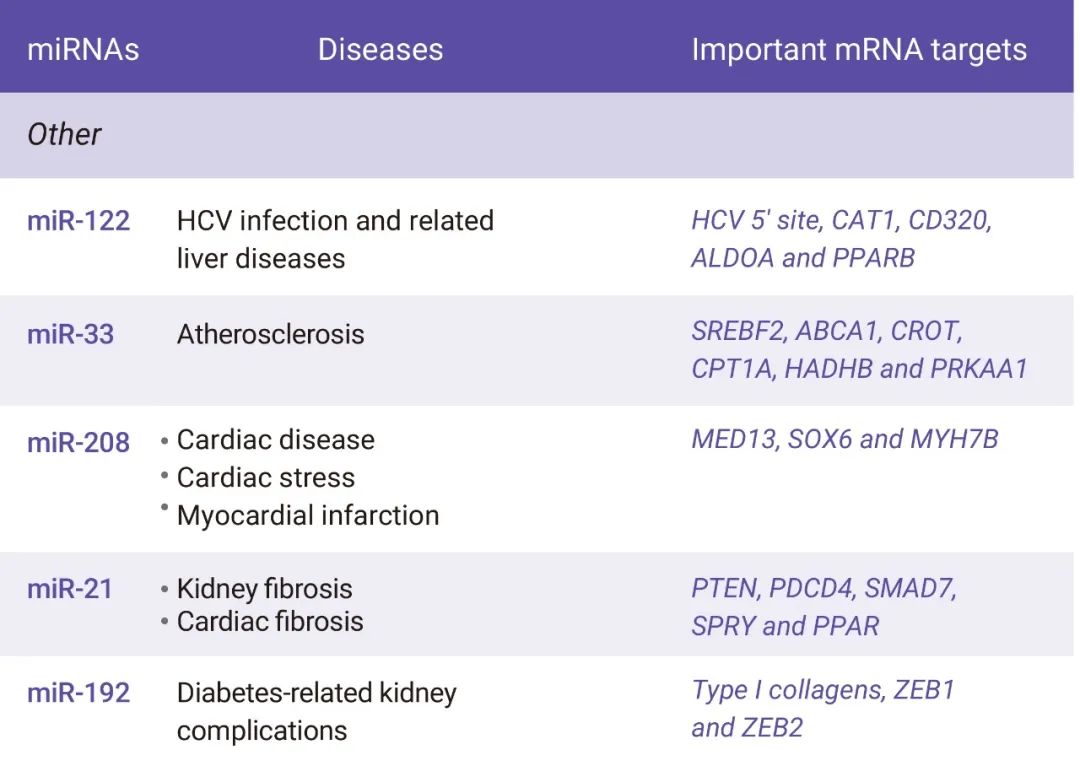
在细胞质中介导基因沉默是 miRNA 的 “看家本领”。由于 miRNA 的低互补性, miRNA 常常与靶基因不完全配对,所以,它们可同时调控多个靶基因的表达[4] (图 2)。据估计, miRNA 参与了多种关键的生物和细胞过程,如凋亡、分化、发育,增殖、代谢及信号转导通路等,调控人类基因组中多达 30% 的蛋白质编码基因[5]。此外,miRNA 失调常与癌症、糖尿病以及心血管疾病等多种人类疾病相关[3]。随着对 miRNA 的深入研究,人们发现 miRNA 能够以 miRNA 模拟物 (miRNA mimics)、 miRNA 抑制剂 (antimiRs) 或其他形式来靶向在疾病状态下发生改变的多个 mRNA,用于多种疾病治疗 (图 2)[3,6]。
左右滑动
图 2. 癌症和其他疾病中的 miRNA[3]
miRNA 诱导基因沉默
我们知道,miRNA 参与多种疾病的调控,随着对 miRNA 的深入探究。研究人员们发现靶向 miRNA 大致有 2 种策略:1. 通过外源性补充 miRNA 以恢复其在疾病环境中的表达; 2. 通过抑制或阻断 miRNA 来降低 miRNA 的作用 。
■ 策略一:miRNA 模拟物补充 miRNA
我们知道,
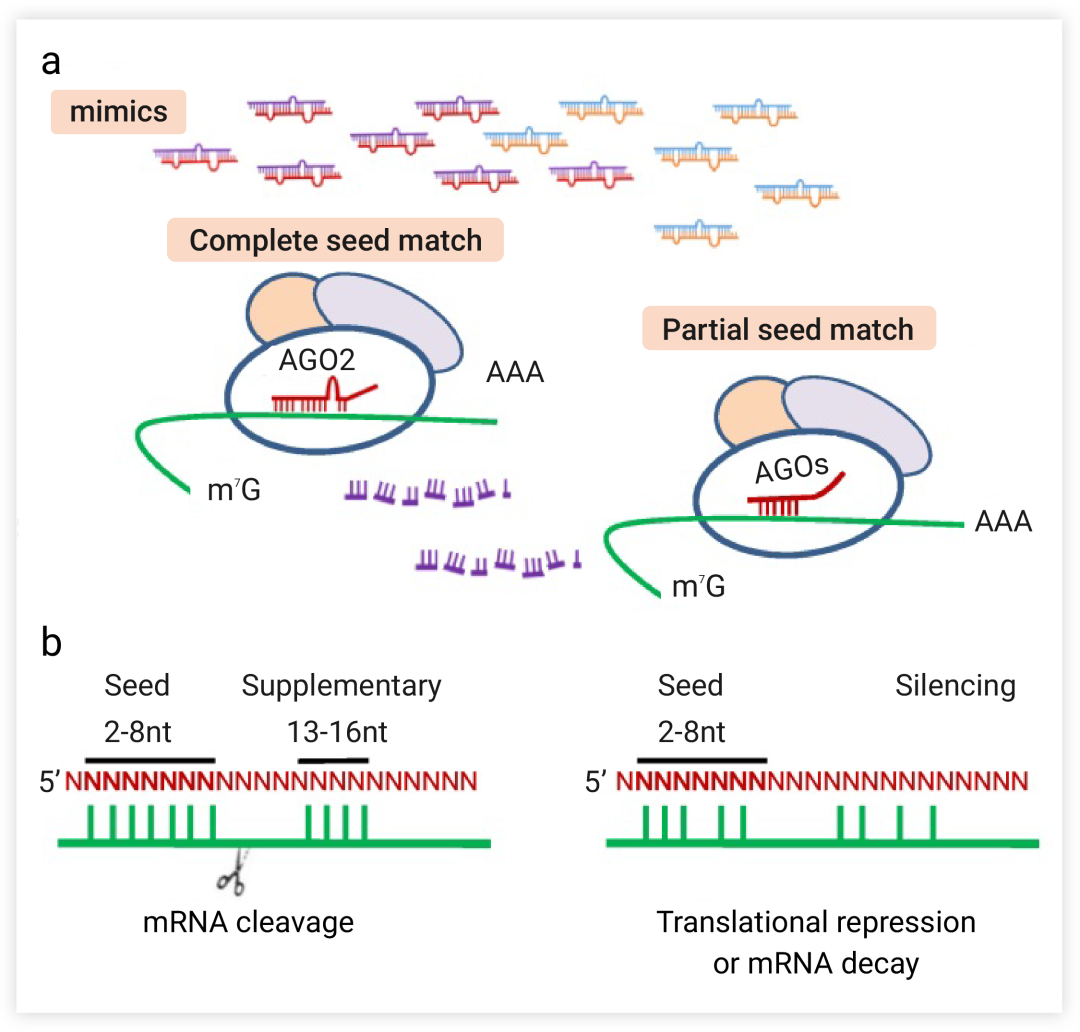
miRNA 可通过指导 RISC 在转录后水平下调基因的表达,从而实现 mRNA 的降解或翻译抑制。那么我们就先来说一说 miRNA 模拟物,作为一种合成的外源性双链寡核苷酸,其具有与内源性 miRNA 相同的序列。miRNA 模拟物主要有两种结合模式:
模式 1:通常情况下,miRNA 成熟后形成 miRISC 复合体,与靶基因 mRNA 的3 '端非编码区结合,通过 miRNA 的种子序列 (seed region) (5’端第 2-8 位的核苷酸序列)识别并结合靶基因 mRNA 上的特异性结合位点,
完全互补匹配的 miRNA 与 AGO2 相互作用,具有核酸酶活性的 RISC 复合体能将靶基因的 mRNA 切割降解,从而抑制靶基因的表达 (图 3a,左)。
模式 2:但大多数动物 miRNA 与 mRNA 的配对是不完美的,存在错配和凸起结合,
miRNA 与靶 mRNA 种子序列部分互补,与 AGO1、3、4 相互作用
。这种情况下,便会
导致 mRNA 的翻译过程被抑制[1] (图 3a,右)。
图 3. miRNA mimics 诱导基因沉默[1]
a. miRNA mimics 的两种结合模式:完全互补的 miRNA 与 AGO2 相互作用 (左);部分种子匹配的 miRNA 与 AGO1、3、4 相互作用 (右);miRNA 模拟物引导链 (红色), 乘客链 (紫色); 内源性 miRNA 引导链 (蓝色) 和乘客链 (橙色); b. 结合模式导向的靶识别导致靶切割或翻译抑制和 mRNA 衰变。
■ 策略二:阻断 miRNA 功能
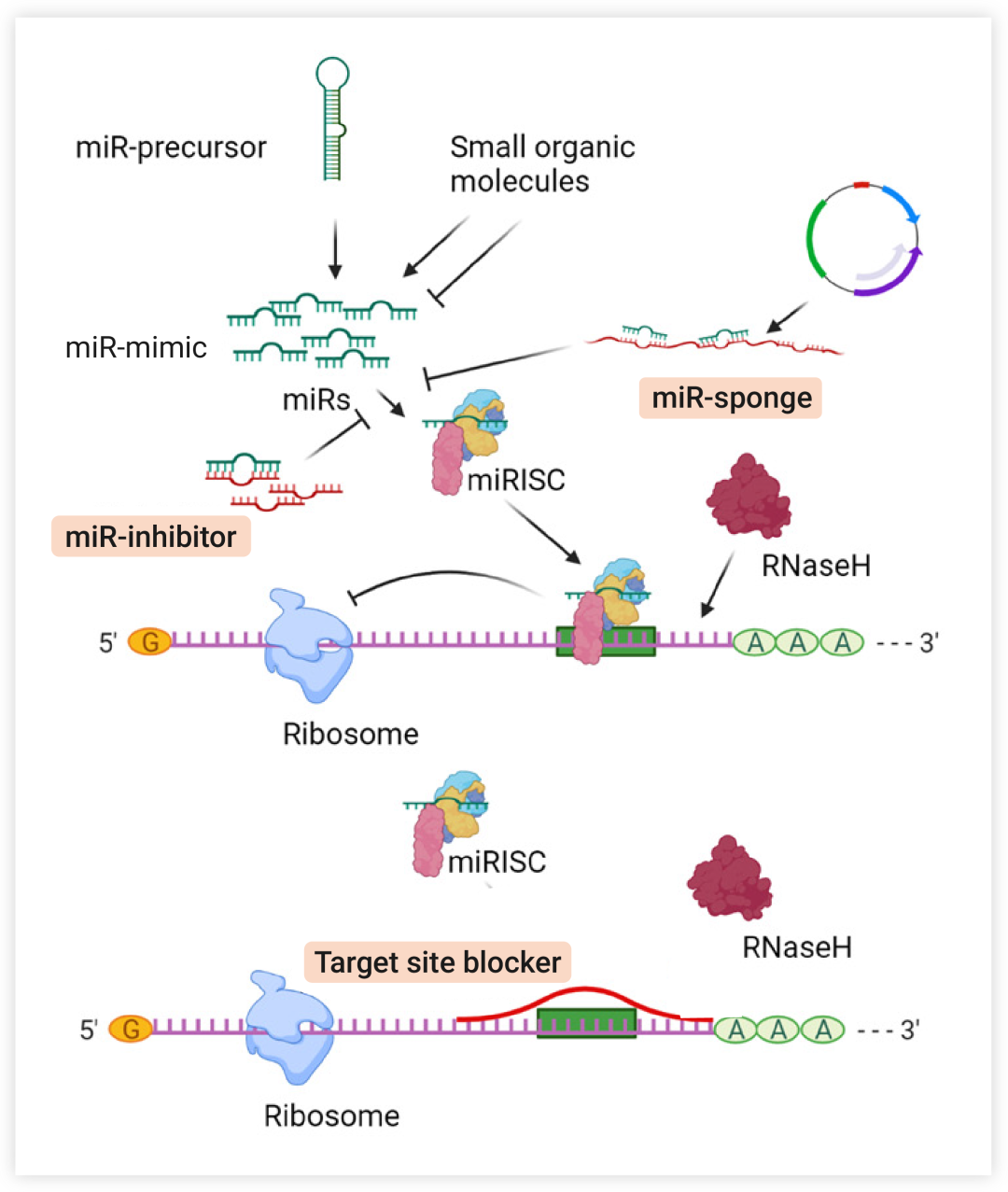
除了补充外源性 miRNA 沉默靶基因外,阻断 miRNA 功能也是靶向 miRNA 的策略之一。主要是通过 miRNA 抑制剂 (antimiRs)、miRNA “阻断剂” (blockmirs) 或者 miRNA “海绵” (miRNA sponges) 来降低 miRNA 的表达 (图 4)。 miRNA 抑制剂一般是指经过化学修饰的短链反义寡核苷酸
(antisense oligodeoxynucleotide, ASO)
,miRNA 抑制剂直接与成熟 miRNA 完全互补配对,可阻断 miRNA 和靶基因的结合。
miRNA 阻断剂与 mRNA 的某些特异性结合位点结合,通过占据结合位点来阻断正常 miRNA 与靶基因 mRNA 的结合, 但这些 “阻断剂” 不影响 miRNA 与其他不具有该特异性结合位点的靶基因结合[7]。
miRNA 海绵通常需要通过病毒载体转染然后才能转录出来。并且 miRNA 海绵同时含有众多 miRNA 结合位点,可以吸引目标 miRNA 与之配对结合,可作为 miRNA 结合的竞争性抑制剂[7]。
图 4. 调控 miRNA 的一般策略[7]
miRNA 抑制剂或 miRNA 海绵可隔离 miRNA 并降低其用于 miRISC 装载的可用性; 靶位点阻断剂阻断 miRNA 与特定靶 mRNA 的结合位点, 导致 mRNA 特异性抑制 miRNA 效应。
■ 实例:miRNA-29b 模拟物抑制肿瘤生长
由于 miRNA 模拟物可模拟内源性 miRNA 的活性,因此,可通过 miRNA 模拟物人为补充外源性 miRNA,增加 miRNA 的总体水平,以恢复相关基因在疾病环境中的表达。例如,在前列腺癌中,miRNA-29b 可靶向沉默 AKT2,从而增加促凋亡基因Bim 的表达,进而抑制肿瘤生长并诱导细胞凋亡 (AKT2 是 miR-29b 的靶点,可抑制促凋亡基因 Bim 的转录[8])。作者通过转染 miR-29b-3p 模拟物,增加 PC3 细胞中miR-29b-3p 的表达,结果发现 miR-29b 过表达后,促凋亡基因 BCL2L11 基因 (Bim) 显著上调 (图 5a-b);前列腺癌细胞增殖呈时间依赖性下降,死亡细胞数量显著增加 (图 5c-d);miR-29b 过表达也抑制了前列腺异种移植瘤生长[9](图 5e-f)。
图 5. miRNA-29b 过表达在前列腺癌中的影响[9]
a-b:用 Bim 特异性抗体 Western blot 分析 (a) PC3 细胞的细胞裂解液和 (b) 用 mimic miR-29b 处理的异种移植瘤;c:PC3 细胞转染对照或模拟 miR-29b, 在 0、24、48、72 h, 用台盼蓝染色细胞,用血细胞计计数活细胞数量。数据以三个独立实验的平均值±标准差表示;d: 用 Calcein AM (绿色为活细胞)和 Ethidium homodimer-1 (红色为死细胞) 染色对照或转染miR-29b 的 PC3 细胞, 通过荧光显微镜定量活细胞和死细胞。箭头表示死亡细胞。右图显示死亡细胞的定量,由五个随机区域计算; e: 对照和实验小鼠的代表性肿瘤; f: 每周监测 2 次肿瘤体积,以平均值表示。
类似的,miRNA-34a 可通过下调 Bcl-2 和 SIRT1 抑制乳腺癌的增殖和迁移;miR-520 可抑制 Ephrin 信号传导,从而显著减少卵巢癌小鼠模型的肿瘤[3]。此外, miRNA 前体也可以增加成熟 miRNA 的水平[10]。不过,也有研究表明 miRNA 模拟物在靶基因抑制中的作用可能依赖于它的浓度以及细胞类型。同时,在不合适的浓度下使用 miRNA 模拟物可能会产生相反的效果[11]。 此外,阻断 miRNA 的功能同样会对疾病产生调控作用,据报道 miR-21 可在受损肾脏中放大损伤和纤维化,由于 miR-21 可直接靶向过氧化物酶体增殖剂激活受体 α (Peroxisome proliferator activated receptor-α, PPARα),从而抑制 PPARα 的转录和表达,这破坏了 PPARα 负调活性氧的过程,造成活性氧的积累,最终导致肾细胞凋亡。特异性沉默 miR-21 可以有效逆转 miR-21 在肾损伤中的有害作用[12]。因此,调控 miRNA 有可能成为一种新的治疗策略。
■ 小结
MiRNA 是一种短的单链 RNA (约 22 个核苷酸,pre-miRNA 是发夹状单链 RNA),其可以与 AGO 蛋白结合形成 RISC 复合体,触发 RNA 干扰,调节靶基因的表达。此外,miRNA 参与了多种关键的生物和细胞过程,包括凋亡、分化、发育,增殖、代谢及信号转导通路等,与多种人类疾病相关,如癌症、糖尿病以及心血管疾病等。





 楼主
楼主