登陆查看更多优秀资源帖,与同道便捷交流讨论
您需要 登录 才可以下载或查看,没有帐号?立即注册
x
铁死亡是一种铁依赖性的,区别于细胞凋亡、细胞自噬的细胞程序性死亡 (programmed cell death,PCD) 方式,依赖于铁介导的氧化损伤,铁积累的增加,自由基的产生,脂肪酸供应与脂质过氧化物增加是诱导铁死亡的关键。
铁死亡的调控机制
关于铁死亡的调控,主要受 GSH/GPX4 途径的调节;铁代谢以及脂质代谢的相关通路也发挥着重要作用。
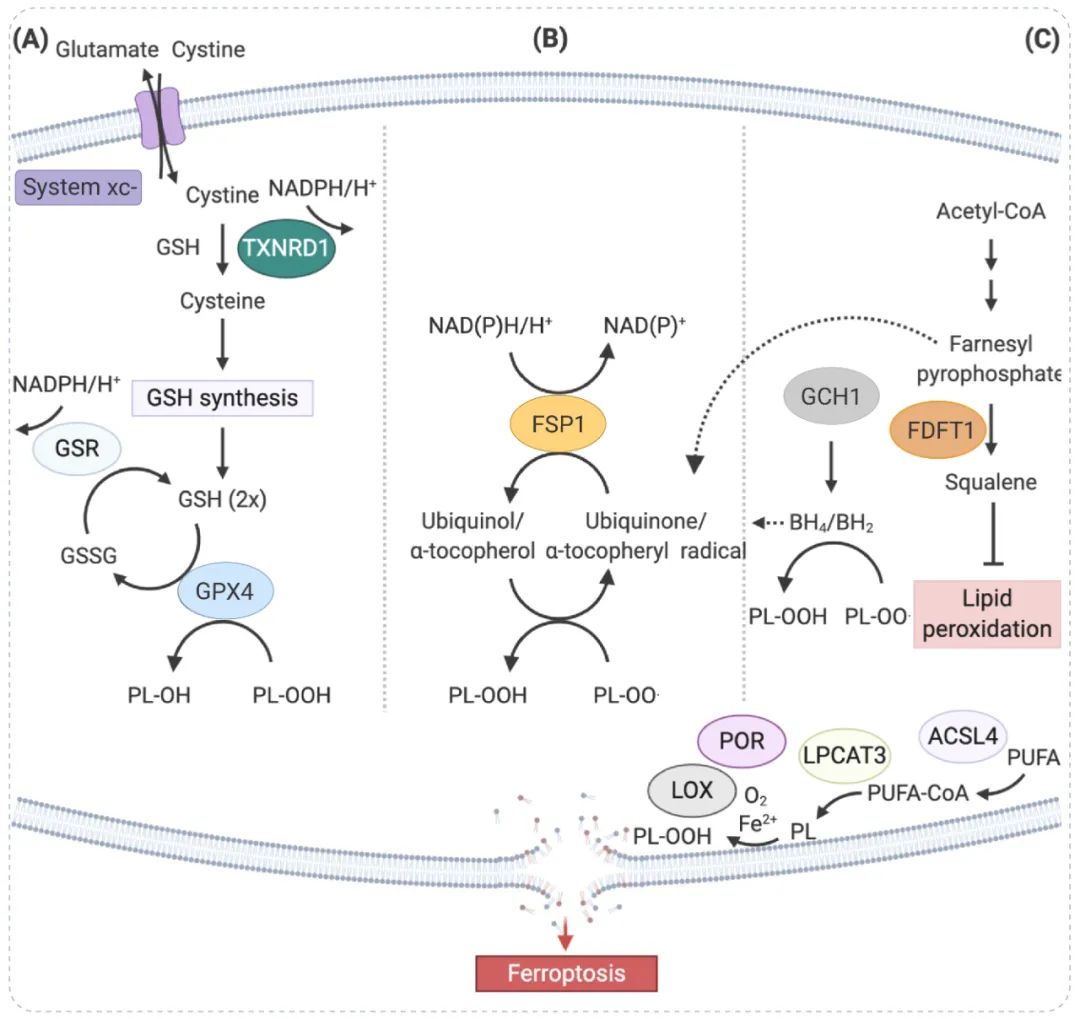
图 1. GPX4 相关信号通路[1] System Xc- 是一种广泛分布在磷脂双分子层中的重要的抗氧化体系,由两个亚基 SLC7A11 和 SLC3A2 组成的异二聚体。胱氨酸和谷氨酸通过 Xc-系统 以 1:1 的比例进出细胞交换,被吸收的胱氨酸 (Cystine) 在细胞中被还原为半胱氨酸 (Cysteine),并参与 GSH 的合成。
GPX4 是催化哺乳动物细胞中磷脂氢过氧化物 (PLOOH) 还原的主要酶。GPX4 将谷胱甘肽 (GSH) 转化为氧化型谷胱甘肽 (GSSG),并将细胞毒性脂质过氧化物 (PL-OOH) 还原为相应的醇 (PL-OH)。因此,GPX4 活性的抑制可导致脂质过氧化物的积累。GPX4 表达下调的细胞对铁死亡更敏感,而 GPX4 表达上调则抑制铁死亡。GSH 在谷胱甘肽过氧化物酶 (GPX) 的作用下减少 ROS 和活性氮。
此外,甲羟戊酸 (MVA) 途径通过调节硒代半胱氨酸 tRNA 的成熟来影响 GPX4 的合成,从而调节铁死亡的发生。
小贴士: Erastin 作为最经典的铁死亡诱导剂,可抑制胱氨酸输入,影响 GSH 的合成,导致 GPX 活性降低,细胞抗氧化能力下降,脂质 ROS 积累, 最终发生氧化损伤和铁死亡。
常用的铁死亡诱导剂 RSL3 直接作用于 GPX4 并抑制其活性,从而降低细胞的抗氧化能力并积累 ROS,导致铁死亡。
图 2. 脂代谢与铁死亡相关通路[2]
铁依赖性脂质 ROS 积累与所有途径的铁死亡有关。不受限制的脂质过氧化是铁死亡的标志。 游离多不饱和脂肪酸是合成脂质信号转导介质的底物,但它们必须被酯化成膜磷脂并被氧化才能传递铁死亡信号。脂氧合酶 (LOX) 和细胞色素 P450 氧化还原酶 (POR) 通过脂质的双氧合启动脂质过氧化。研究表明,Phosphatidyl ethanolamine 是诱导细胞铁死亡的关键磷脂。酰基辅酶 A 合成酶长链家族成员 4 (ACSL4) 和溶血磷脂酰胆碱酰基转移酶 3 (LPCAT3) 参与Phosphatidyl ethanolamine 的生物合成和重构,激活多不饱和脂肪酸并影响其跨膜特性。 因此,降低 ACSL4 和 LPCAT3 的表达可减少细胞内脂质过氧化物底物的积累,从而抑制铁死亡。
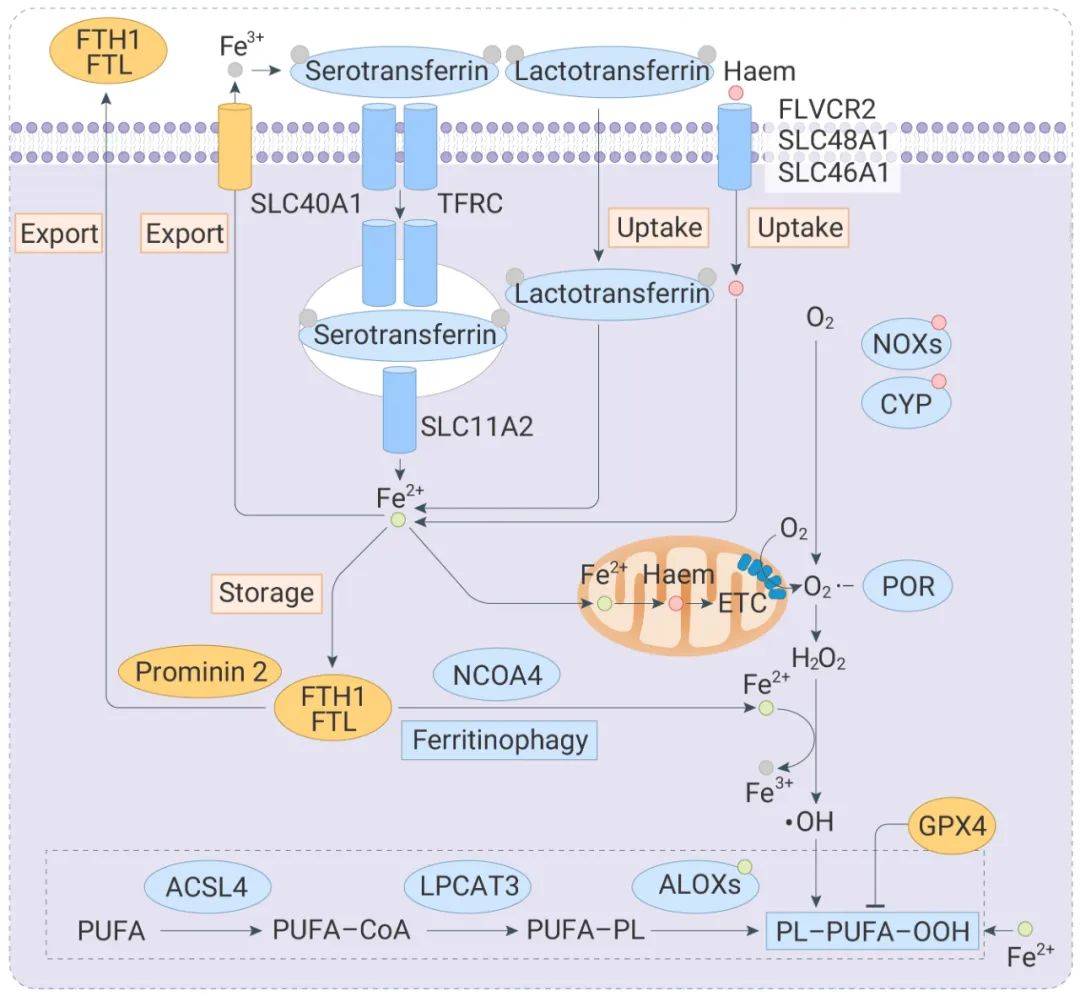
铁积累 (如增加铁吸收、减少铁储存和限制铁外流)可以促进铁死亡 (具体可见: 铁死亡是什么,如何检测?您要的“一文通”来了!)。过量的铁通过至少两种机制促进随后的脂质过氧化:通过铁依赖性 Fenton 反应产生活性氧 (ROS) 以及激活含铁酶。
铁死亡如何检测呢?
铁死亡的关键指标铁死亡是由铁依赖的脂质过氧化驱动的,因此在铁死亡期间检测这种脂质过氧化是必要的。此外,线粒体在铁死亡期间通常表现出萎缩、致密的形态(因此可以通过检测线粒体来判断铁死亡)。此外,检测特定的基因表达变化:例如铁死亡的细胞中 TfR1 的上调及其重定位于质膜。下面小 M 来给大家介绍几种常用的铁死亡检测方式~
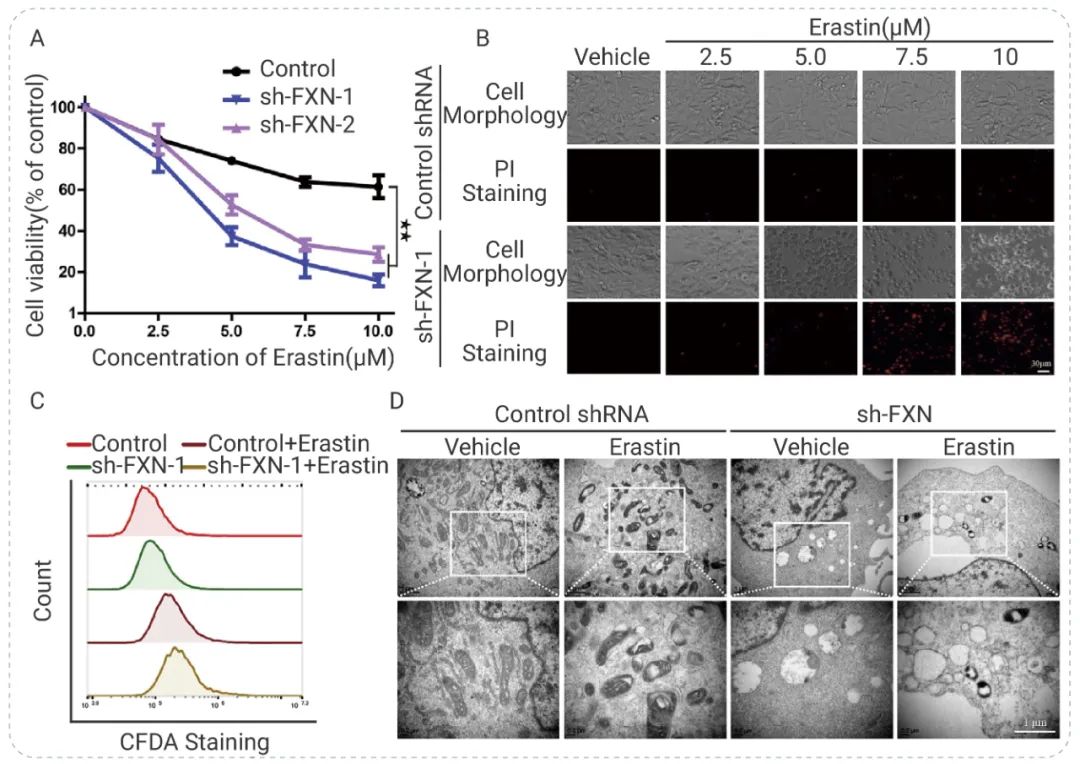
铁死亡会导致细胞死亡。因此一般在铁死亡的研究中,检测细胞活力是常见方法,同时也会通过染色等方法来观察细胞膜通透性,线粒体形态等。 如在 Identification of Frataxin as a regulator of ferroptosis 一文中,作者研究抑制 Frataxin (FXN) 是否会促进铁死亡。用 Erastin 同时处理 FXN 敲低和对照细胞,12 小时后用 CCK8 法检测细胞活力,结果表明抑制 FXN 表达显著增强了 Erastin 诱导的细胞死亡。同时碘化丙啶 (PI) 和 CFDA-SE 染色结果以及用透射电镜观察到的线粒体碎裂、空泡化和嵴增大都表明 FXN 耗竭协同 Erastin 诱导铁死亡[3]。图 3. 抑制 FXN 对 Erastin 诱导的 HT-1080 细胞铁死亡的影响[3] A, 细胞暴露于不同浓度的 Erastin,检测细胞活力;B, Erastin 处理细胞 12 小时后用荧光显微镜观察 PI 阳性细胞;C, Erastin 培养细胞 3 天,用 CFDA-SE (5 μM) 染色,然后进行流式检测;D, 透射电子显微镜 (TEM) 检测线粒体的形态变化
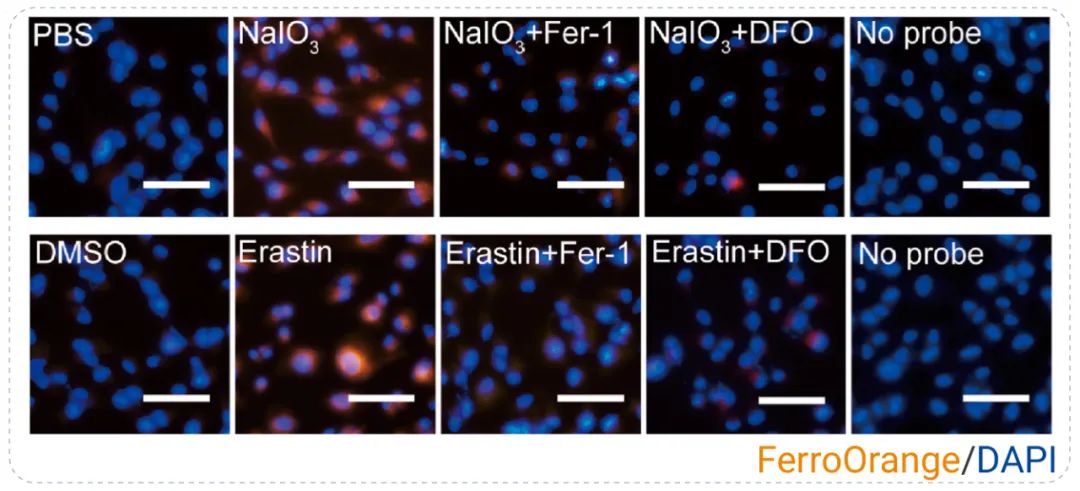
细胞铁积累是铁死亡的典型标志之一,亚铁离子积累可以特异性地增加氧化应激水平。例如,在研究氧化应激介导的视网膜色素上皮 (RPE) 变性的潜在机制时,通过用 FerroOrange 染色细胞 (亚铁离子探针),结果表明 NaIO3 和 Erastin 处理的细胞积累了过多的亚铁离子,说明诱导了铁死亡,而 Fer-1 或 DFO 预孵育可以抑制亚铁离子的积累,如图 4 所示。结果表明铁死亡是氧化应激介导的 RPE 变性的主要病理过程[5]。
图 4. 通过 FerroOrange 染色评估亚铁离子水平[4]
ROS 和脂质 ROS 在铁死亡中起关键作用,文献报道增加的超氧化物歧化酶 (SOD) 可抑制体内的 ROS 水平。细胞内会由于铁的积累而抑制抗氧化系统,铁可能直接通过 Fenton 反应产生过量的 ROS,从而增加氧化损伤。因此 ROS 检测也是常用方法 (活性氧检测探针: ROS 探针大赛,你要的检测方法都在这里!)
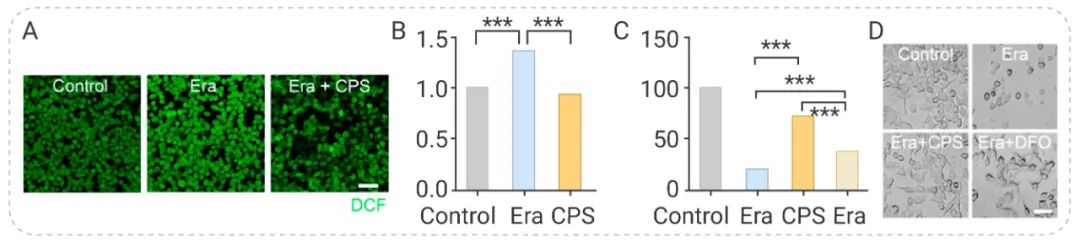
Lin Li 在研究中发现羧基修饰的 (CPS) 可以增加细胞内 SOD 水平,因此推测 CPS 可能抑制铁死亡。实验结果表明,CPS 抑制了 RAW 264.7 中 Erastin 诱导的 ROS 升高 (图 5a, b),并且比去铁胺具有更强的抗铁死亡作用 (图 5c, d)[5]。
图 5. CPS 有效抑制 RAW 264.7 细胞的铁死亡[5]
A-B, 在有无 CPS 条件下 Erastin 对 ROS 水平的影响;C-D, 在有无 CPS 或 DFO 条件下Erastin 对细胞活力的影响
除了 ROS 检测,铁死亡的脂质过氧化的指标还有谷胱甘肽 (GSH) 合成的变化、脂质过氧化物 (MDA, LPO) 水平的变化。
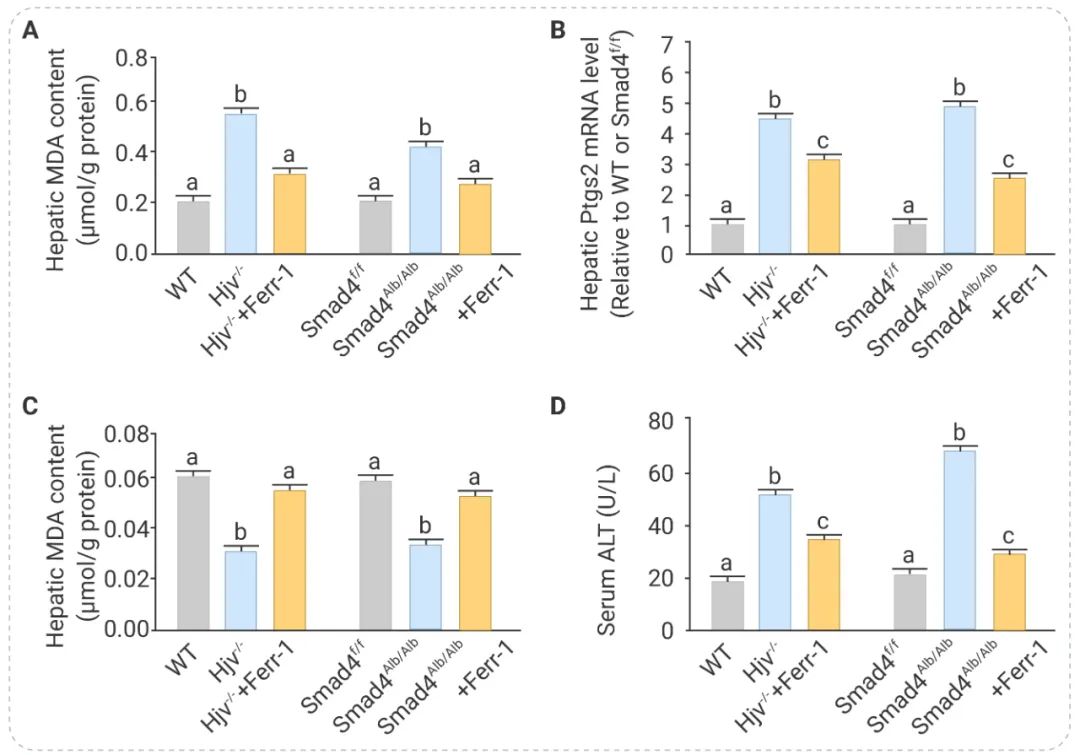
在遗传性血色素沉着症 (HH,一种铁超负荷疾病) 与铁死亡的关系研究中,为了证实铁死亡在 HH 相关肝损伤中的作用,作者用铁死亡抑制剂 Fer-1 治疗 Hjv–/– 小鼠 (经典的 HH 小鼠模型) 和 Smad4 Alb/Alb 小鼠 (HH 样小鼠模型) 三周。如图 6 所示,与对照组小鼠相比,经 Fer-1 处理的小鼠肝脏 MDA 水平显著降低,NADPH 水平增加。此外,Fer-1 可以降低小鼠血清 ALT 水平胶原蛋白沉积。结果表明,铁超载会诱导 HH 小鼠的铁死亡[6]。 图 6. 铁死亡抑制剂 Fer-1 可减轻 HH 小鼠中铁过载引起的肝损伤[6] 用或不用 Fer-1 治疗的野生型、Hjv–/–、Smad4Flox/flox 和 Smad4Alb/Alb 小鼠中, A, 肝脏 MDA 含量;B, 肝脏 Ptgs2 mRNA 水平;C, 肝脏 NADPH 含量;D, 血清 ALT 水平
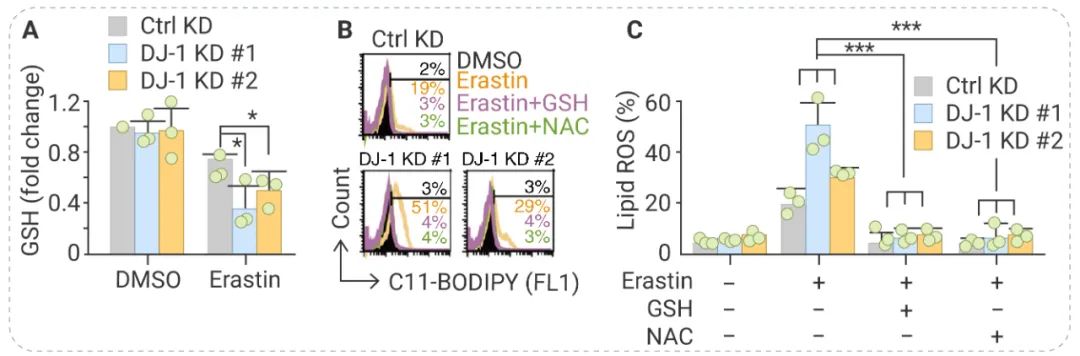
在 DJ-1 suppresses ferroptosis through preserving the activity of S-adenosyl homocysteine hydrolase 一文中,作者探究了 DJ-1 负调节铁死亡的机制。研究发现,用 RNA 干扰抑制 DJ-1 表达可以与 Erastin 协同抑制细胞内 GSH 水平 (图 7A)。同时,添加外源性 GSH 或 N-乙酰半胱氨酸 (NAC) 可以逆转 Erastin 诱导的脂质 ROS 积累 (图 7B) 和细胞死亡 (图 7C)。因此,作者推测 DJ-1 可能会影响 GSH 的合成从而抑制铁死亡[7]。 图 7. DJ-1 通过抑制 GSH 水平负调节铁死亡[7] A, Erastin 处理DJ-1 KD H1299 细胞 6 h 后 GSH 水平; B, 在有无 GSH 或 NAC 条件下细胞用 Erastin 处理细胞 12 h 后脂质 ROS 水平;C, 用Erastin 处理细胞 36 h 后,测定细胞活力
总结: 铁死亡受细胞内信号通路的严密调节,既包括铁稳态的调节通路,更有铁代谢以及脂质代谢相关通路等。 但是各个通路相辅相成,互相影响。调控途径不同, 检测的指标也十分多样化,小伙伴们在实验过程中,检测方法可要多打 “组合拳”!
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务
参考文献 1. Jiang X, et, al. Ferroptosis: mechanisms, biology and role in disease. Nat Rev Mol Cell Biol. 2021 Apr;22(4):266-282. 2. Chen X, Kroemer G, Tang D, et, al. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021 May;18(5):280-296. 3. Du J, Zhou Y, Li Y, Tong X, Wang Y, et, al. Identification of Frataxin as a regulator of ferroptosis. Redox Biol. 2020 May;32:101483. 4. Tang Z, Ju Y, Dai X, Zhang J, Gu P ,et, al. HO-1-mediated ferroptosis as a target for protection against retinal pigment epithelium degeneration. Redox Biol. 2021 Jul;43:101971. 5. Wang H, et, al. Characterization of ferroptosis in murine models of hemochromatosis. Hepatology. 2017 Aug;66(2):449-465. 6. Li L, et, al. Polystyrene Nanoparticles Reduced ROS and Inhibited Ferroptosis by Triggering Lysosome Stress and TFEB Nucleus Translocation in a Size-Dependent Manner. Nano Lett. 2019 Nov 13;19(11):7781-7792. 7. Cao J, et, al. DJ-1 suppresses ferroptosis through preserving the activity of S-adenosyl homocysteine hydrolase. Nat Commun. 2020 Mar 6;11(1):1251.
| 




 楼主
楼主