登陆查看更多优秀资源帖,与同道便捷交流讨论
您需要 登录 才可以下载或查看,没有帐号?立即注册
x
话说线粒体自噬,可谓是大家眼中的“老大哥”了。往期小 M 为大家介绍过自噬的分类,底物以及检测 (详见往期推文:Hello~自噬;还能这么玩儿?跟大牛解锁自噬检测),今天小 M 带大家进一步了解线粒体自噬的经典机制以及检测方法~线粒体自噬:细胞器的选择性自噬
线粒体,作为一种双层膜半自主细胞器,它是细胞中制造能量的结构,是细胞有氧呼吸和利用氧化磷酸化产生 ATP 的场所的主要场所。其 “大佬” 地位毋庸置疑,但它的功能状态与线粒体膜电位、线粒体膜通道、线粒体 Ca2+ 浓度、呼吸链复合体活性、活性氧生成以及 DNA 突变密切相关。线粒体质量控制通过蛋白质平衡、线粒体自噬、动力学和生物发生等方式的协调来维持线粒体的完整性和功能。
■ 什么时候发生线粒体自噬?
线粒体自噬 (Mitochondrial autophagy, mitophagy) 作为一种重要线粒体质量控制机制,在活性氧 (ROS) 胁迫等应激作用下,会导致线粒体 DNA (Mitochondrial DNA, mtDNA) 突变逐渐累积,还会使细胞内线粒体膜电位降低和去极化损伤,并最终导致细胞死亡[1][2]。严峻的生存形势下,线粒体只好 “大开杀戒”,为了维持线粒体和细胞稳态,防止受损线粒体损伤细胞,细胞通过选择性地包裹和降解细胞内受损或功能障碍的线粒体--即线粒体自噬。
既然被吃已然是定局,那么受损线粒体是如何被 “干掉” 的?这一过程与巨自噬有着很大的相似性,但它更像是细胞器的选择性自噬去除。
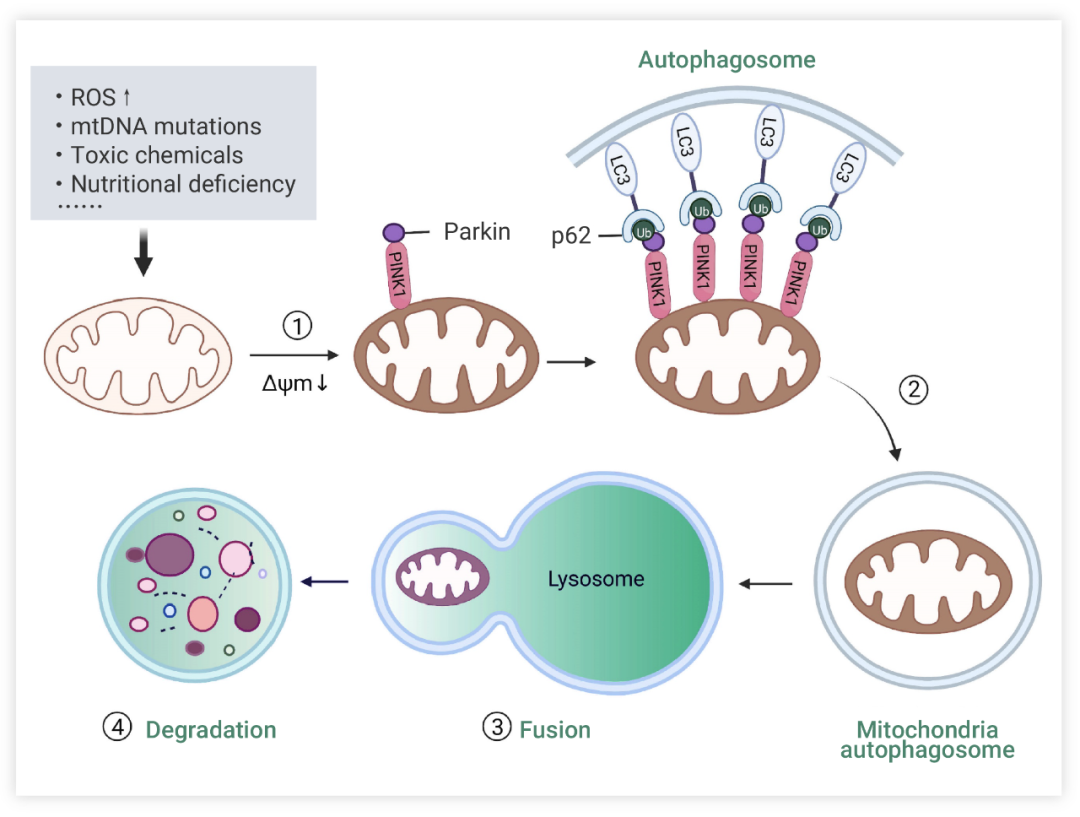
线粒体自噬主要有以下 4 个关键步骤:1) 受损线粒体去极化,失去膜电位。2) 线粒体被自噬体包裹形成线粒体自噬体。3) 线粒体自噬体与溶酶体融合。4) 线粒体内容物被溶酶体降解。溶酶体或液泡酸性水解酶流入自噬体降解受损线粒体。
图 1. 线粒体自噬的关键步骤[1]
线粒体自噬途径的相关机制
线粒体自噬的机制通常分为两类: 泛素依赖途径和非泛素依赖途径。
■ 泛素依赖性途径
花开两朵,各表一枝。首先,我们来看一看泛素依赖性途径,顾名思义,依赖于线粒体表面蛋白的广泛泛素化来促进线粒体自噬。目前研究最广泛的便是 PINK1/Parkin 通路。在这一机制中,PTEN 诱导的激酶 1 (PTEN induced kinase 1, PINK1) 堪称线粒体自噬的得力助手。当线粒体膜电位 (Mitochondrial membrane potential, MMP, ΔΨm) 受损时,PINK1 进入线粒体内膜的途径受阻,导致 PINK1 在线粒体外膜的胞质面上稳定聚集。同时,这会募集并激活 Parkin,Parkin 蛋白酶的空间构象发生改变,转化为活化的 E3 泛素连接酶,然后泛素化线粒体上的蛋白质[3]。PINK1 与 Parkin 相互作用,共同调控线粒体自噬过程以维持线粒体质量 (图 2)。此外,除了 PINK1-Parkin 通路之外,还有非 Parkin 依赖性的泛素依赖性通路。也就是说,PINK1 还可以通过泛素磷酸化直接招募自噬受体蛋白 (如 NIX、BNIP3 和 FUNDC1) 到线粒体,受体蛋白募集 LC3,这使得自噬体能够吞噬线粒体。
 图 2. 线粒体自噬机制概述[1]
线粒体自噬的 Ub 依赖途径 (PINK1/Parkin 通路最为常见) 和 Ub 非依赖性途径
图 2. 线粒体自噬机制概述[1]
线粒体自噬的 Ub 依赖途径 (PINK1/Parkin 通路最为常见) 和 Ub 非依赖性途径
■ 非泛素依赖性途径
尽管据了解 PINK1 和 Parkin 的激活可以触发线粒体自噬,但线粒体自噬也可能通过其他机制进行:相比之下,非泛素依赖途径就略显 “清闲”,线粒体外膜 (OMM)上有许多包含 LC3 相互作用区 (LIR) 区域的蛋白,它们是自噬的受体。它们可以不经泛素化直接与 LC3 结合,从而启动线粒体自噬 (图 2)。
在哺乳动物中,这些受体主要包括 Nip3 样蛋白 X (Nip3-like protein X, NIX) 受体、bcl2 相互作用蛋白 3 (BCL2-interacting protein 3, BNIP3) 受体、FUN14 结构域包含 1 (FUN14 domain containing 1, FUNDC1) 受体
■ 线粒体自噬的相关疾病
正常的线粒体活动对细胞功能至关重要,及时的消除受损线粒体是细胞的自我保护机制。当线粒体自噬发生障碍可触发多种疾病,如:线粒体功能障碍是阿尔茨海默氏症,帕金森氏症,亨廷顿氏症等神经退行性疾病的关键共同因素,PINK1 和 Parkin 的功能缺失突变与家族性帕金森病相关。心肌细胞的活动高度依赖于线粒体的能量供应,线粒体吞噬功能障碍可引起心肌肥厚、心律失常、心源性猝死等心血管疾病[6]。
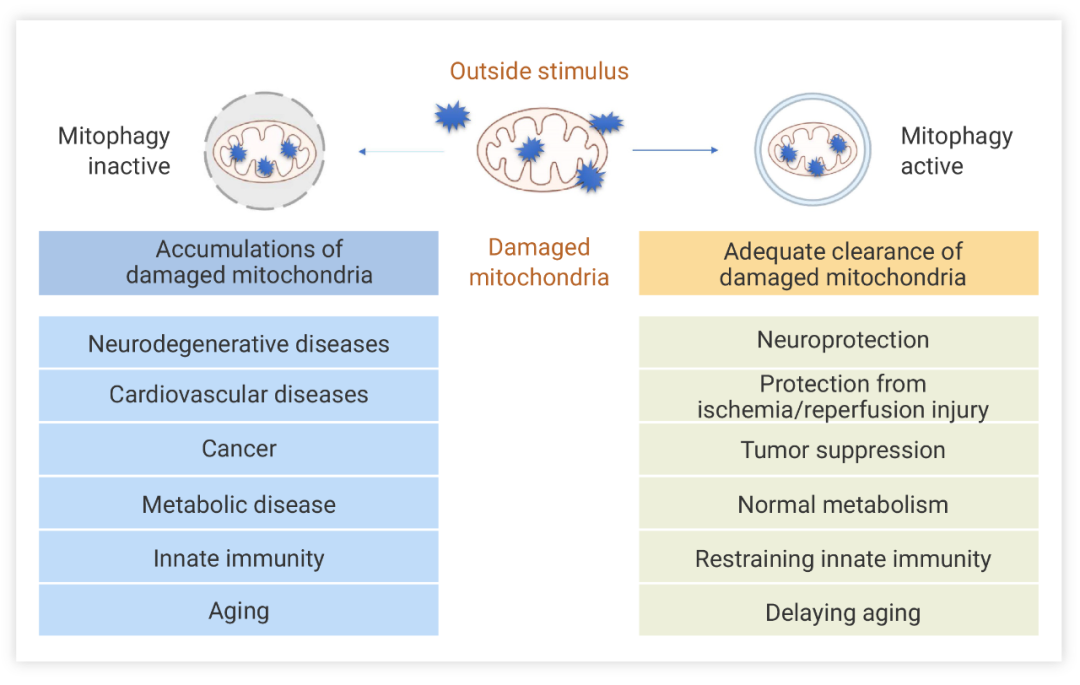
图 2. 线粒体自噬的相关疾病[1]
线粒体自噬,该如何检测?
线粒体自噬是一个复杂的、动态的过程,检测方法也在不断的更新,小 M 以文献为例,盘点常见的线粒体自噬研究方法!
Myoferlin 是一种在多种癌症中过度表达的癌蛋白,已有报道表明 Myoferlin 通过与线粒体动态机制的相互作用对胰腺癌的线粒体适应性有显著贡献[7]。为证明 Myoferlin 在 PDAC 细胞系中对线粒体自噬的影响,研究者使用了 WJ460 处理 PDAC 细胞并进行线粒体形态观察,如图所示 WJ460 处理导致线粒体嵴结构紊乱,甚至出现空白区域 (图 4a) 。研究者还进行了对自噬体和线粒体进行了免疫定位:WJ460 处理的癌细胞中自噬体的数量增加,并且在 BxPC-3、Panc-1 和 PaTu 8988T 细胞系中发现了共定位点 (图 4b)。此外,PDAC 在 WJ460 处理后,线粒体 ROS 丰度显着降低 (图 4c) 。以上表明,在 PDAC 细胞系中,Myoferlin 的靶向抑制引起线粒体能量应激,并引发线粒体自噬。
图 4. WJ460 在 PDAC 细胞诱导线粒体自噬[7]
a. 电镜下观察线粒体状态。b. 免疫荧光共定位。c. 活性氧检测
除上述线粒体形态观察 (透射电镜下线粒体受损情况),ROS 浓度测定线粒体内活性氧的积累,自噬体与线粒体免疫荧光共定位等方法,线粒体自噬的相关追踪探针以及线粒体自噬标志物的 Western 检测以及也是常用的检测线粒体吞噬的方法[8]。
如,在今年发表的A mitochondrial SCF-FBXL4 ubiquitin E3 ligase complex degrades BNIP3 and NIX to restrain mitophagy and prevent mitochondrial disease 一文报道了关于线粒体自噬调控和线粒体疾病的研究。研究发现,在线粒体疾病 MTDPS13 中突变的蛋白 FBXL4 定位在线粒体外膜并发现 FBXL4 负调控线粒体自噬,抑制线粒体自噬的过度激活。作者团队使用 mtKeima 报告基因分析了线粒体吞噬, mtKeima 是一种针对线粒体基质的 ph 敏感蛋白荧光蛋白,Keima 的激发光谱会随 pH 值而变化(图 5a)。短波长在中性环境中的激发,而长波长会酸性环境激发,可用于区分游离线粒体和线粒体溶酶体。
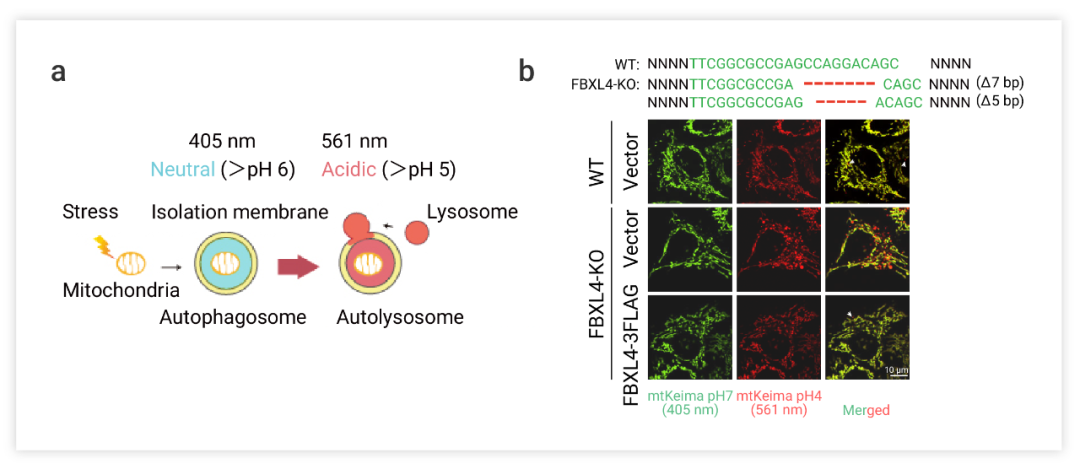 图 5. FBXL4-KO在HeLa细胞诱导线粒体自噬[9]
a. 探针追踪机制的示意图。b. 免疫荧光共定位
图 5. FBXL4-KO在HeLa细胞诱导线粒体自噬[9]
a. 探针追踪机制的示意图。b. 免疫荧光共定位
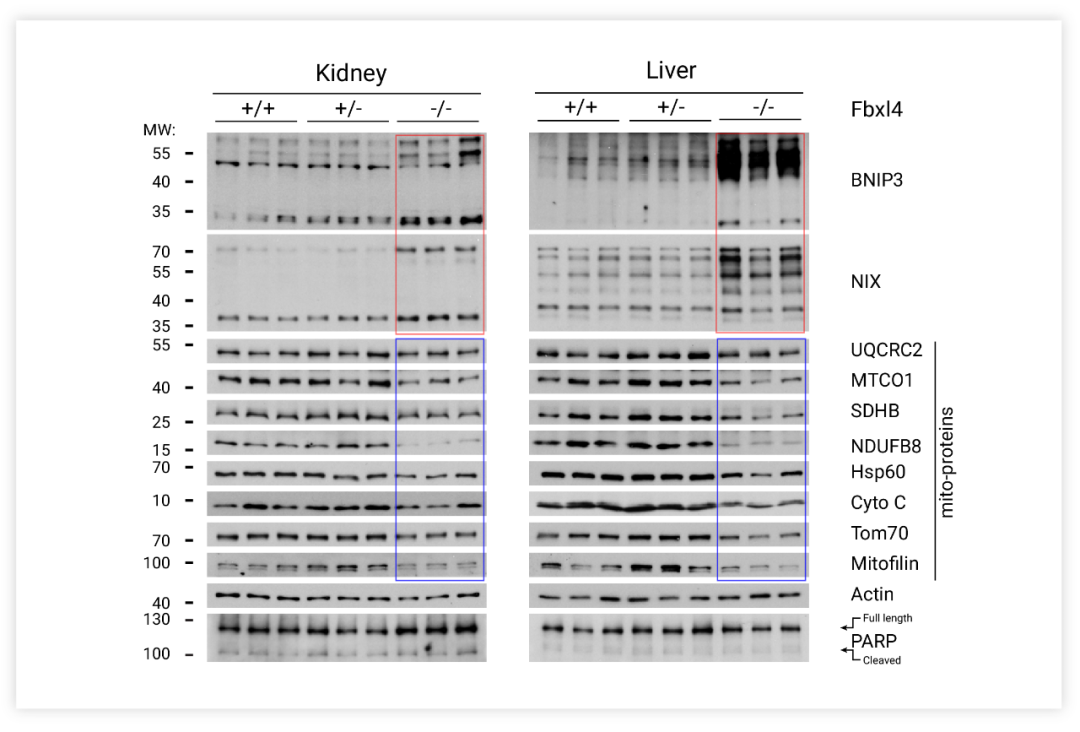
当线粒体被溶酶体吞噬时,Keima 会处在酸性环境,如图在 Merged 图中,黄绿色的 mtKeima 信号标记细胞质内的线粒体,红色的 mtKeima 信号标记溶酶体内的线粒体自噬。FBXL4-KO 导致线粒体数量过少,激活了线粒体自噬。自噬过程中,参与线粒体自噬的蛋白 (PINK1、Parkin、BNIP3、Nix、FUNDC1) 水平在线粒体吞噬被激活后会增加。如图 7 所示, Fbxl4-/-小鼠的组织样本检测出标记物蛋白 BNIP3 和 NIX 的积累、过度线粒体自噬和线粒体数量减少。
图 6. Fbxl4-/- 小鼠组织样本的Western 检测[9]
■ 小结
线粒体自噬可谓是非“吃”不可,方式也是多种多样,除了泛素依赖途径 (PINK1/Parkin 通路为主) 和非泛素依赖途径的经典机制。小M 还给大家介绍了线粒体自噬的一些常见检测方法,希望能帮助到小伙伴们!
MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务
参考文献
[1] Lu Y, et al. Cellular mitophagy: Mechanism, roles in diseases and small molecule pharmacological regulation. Theranostics. 2023;13(2):736-766.
[2] Lemasters JJ. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res. 2005;8(1):3-5.
[3] Riley BE, Lougheed JC, Callaway K, et al. Structure and function of Parkin E3 ubiquitin ligase reveals aspects of RING and HECT ligases. Nat Commun. 2013;4:1982.
[4] Lazarou M, Sliter DA, Kane LA, et al. The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature. 2015;524(7565):309-314.
[5] Vargas JNS, Wang C, Bunker E, et al. Spatiotemporal Control of ULK1 Activation by NDP52 and TBK1 during Selective Autophagy. Mol Cell. 2019;74(2):347-362.e6.
[6] Qiu Y, Wang J, Li H, et al. Emerging views of OPTN (optineurin) function in the autophagic process associated with disease. Autophagy. 2022;18(1):73-85.
[7] Gilles Rademaker, et al. Myoferlin targeting triggers mitophagy and primes ferroptosis in pancreatic cancer cells. Redox Biol. 2022 Jul;53:102324.
[8] Tan HWS, Lu G, Dong H, et al. A degradative to secretory autophagy switch mediates mitochondria clearance in the absence of the mATG8-conjugation machinery. Nat Commun. 2022;13(1):3720. Published 2022 Jun 28.
[9]. Yu Cao, Jing Zheng, Huayun Wan, et al. A mitochondrial SCF-FBXL4 ubiquitin E3 ligase complex degrades BNIP3 and NIX to restrain mitophagy and prevent mitochondrial disease. EMBO J. 2023 Mar 10;e113033.
[10] Han R, Liu Y, Li S, Li XJ, Yang W. PINK1-PRKN mediated mitophagy: differences between in vitro and in vivo models [published online ahead of print, 2022 Nov 3]. Autophagy. 2022;1-10.
[11] Wang X, Winter D, Ashrafi G, et al. PINK1 and Parkin target Miro for phosphorylation and degradation to arrest mitochondrial motility. Cell. 2011;147(4):893-906.
[12] Huang E, Qu D, Huang T, et al. PINK1-mediated phosphorylation of LETM1 regulates mitochondrial calcium transport and protects neurons against mitochondrial stress. Nat Commun. 2017;8(1):1399. Published 2017 Nov 9.
[13] Yang W, Liu Y, Tu Z, et al. CRISPR/Cas9-mediated PINK1 deletion leads to neurodegeneration in rhesus monkeys. Cell Res. 2019;29(4):334-336.
[14] Yang W, Guo X, Tu Z, et al. PINK1 kinase dysfunction triggers neurodegeneration in the primate brain without impacting mitochondrial homeostasis. Protein Cell. 2022;13(1):26-46.
|





 楼主
楼主