登陆查看更多优秀资源帖,与同道便捷交流讨论
您需要 登录 才可以下载或查看,没有帐号?立即注册
x

肠道菌群代谢物,顾名思义,由肠道菌群代谢产生。肠道菌群与宿主的相互作用往往通过肠道菌群代谢物来实现。
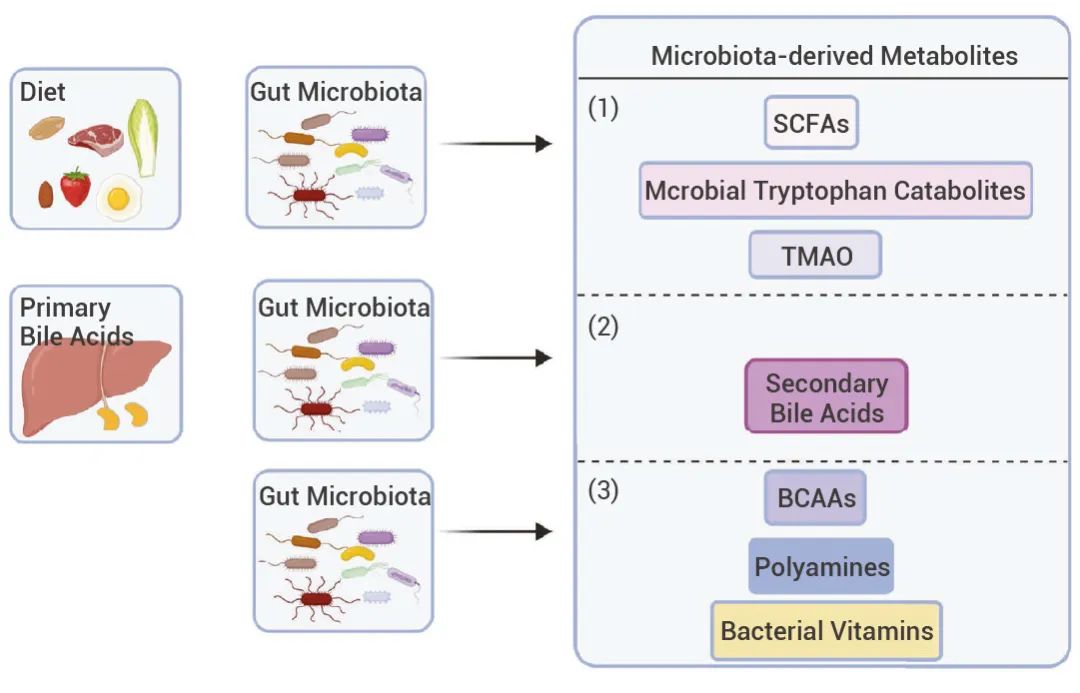
就目前已经鉴定出的肠道微生物群代谢物来说,根据其来源和合成情况,可大致分为三组[1]:
(1)肠道细菌从膳食成分中产生的代谢物;
(2)由宿主产生并经肠道细菌修饰的代谢物;
(3)肠道细菌重新合成的代谢物。图 1. 肠道微生物群衍生代谢物的产生[1]。 ▐膳食产生的代谢物
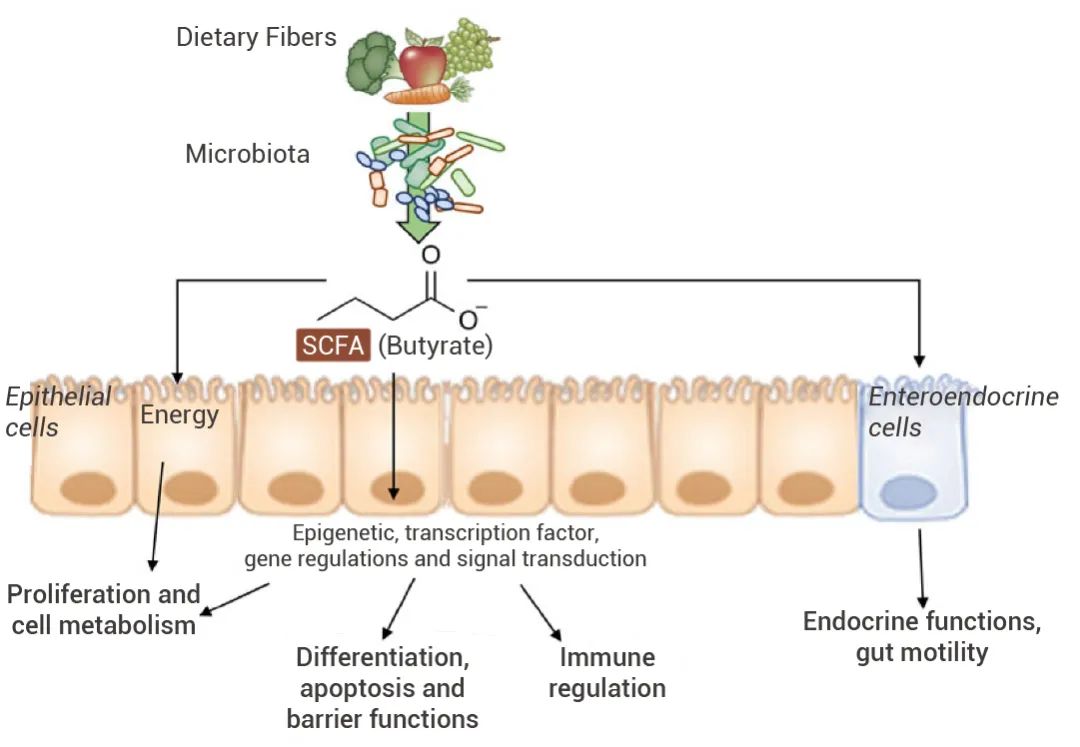
“民以食为天”,食物由口进入胃肠道,未消化的碳水化合物通过肠道微生物群发酵成短链脂肪酸(SCFAs)。 SCFAs 是肠道厌氧菌发酵的主要代谢产物[2],可为不同的组织提供能量,还可调节细胞增殖和分化、激素分泌以及免疫/炎症反应的激活等。如丙酸和丁酸可抑制刺激诱导的粘附分子表达、趋化因子产生,从而抑制单核细胞/巨噬细胞和中性粒细胞募集,具有抗炎作用[2]。
图 2. SCFA 对宿主机能的影响[3]。
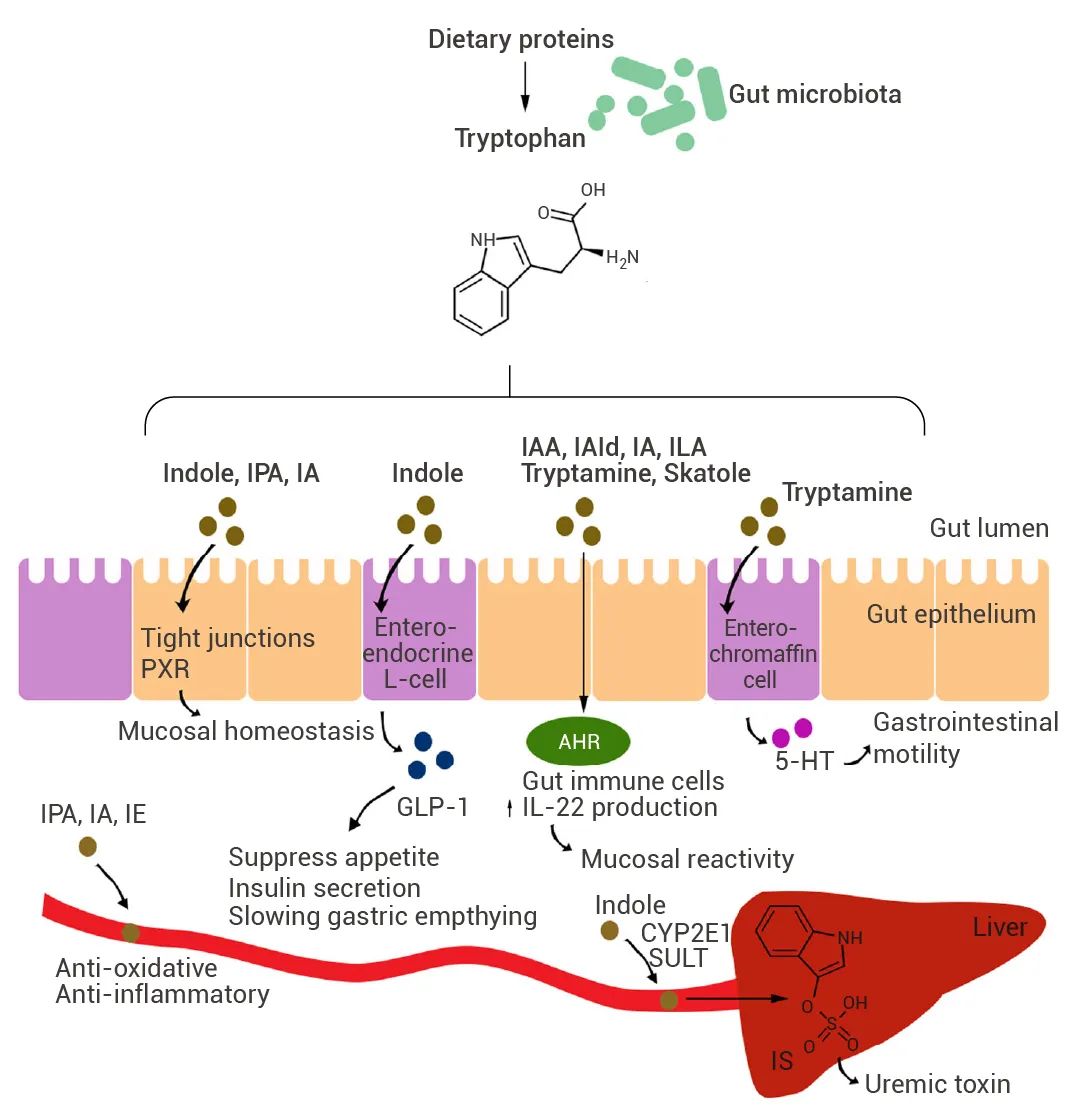
膳食中蛋白质的降解会导致色氨酸的释放,色氨酸可被肠道微生物转化为各种分解代谢物。其中吲哚 (Indole)、IPA 和吲哚丙烯酸 (Indoleacrylic acid, IA) 可通过降低肠道通透性来影响粘膜稳态; 吲哚还可诱导肠内分泌 L 细胞释放 GLP-1。ILA、IAA 、Skatole 等作用于肠道免疫细胞中的 AHR,从而以配体特异性方式改变先天性和适应性免疫反应; Tryptamine 通过诱导释放 5-HT 刺激胃肠蠕动。此外,色氨酸分解代谢物可通过肠上皮吸收并进入血液,其中一些(如 IPA、IE、IA)具有抗氧化和抗炎作用,而硫酸吲哚酚 (IS) 在高浓度下具有细胞毒性作用[4]。 图 3. 微生物色氨酸分解代谢物对宿主生理的作用机制[3]。
而动物性食品中的肉碱、胆碱类化合物可被肠道菌群代谢为三甲胺,随后进入肝脏被黄素单加氧酶氧化产生 TMAO (Trimethylamine-N-oxide, TMAO)[5]。TMAO 是许多慢性疾病的候选危险因素[6]。
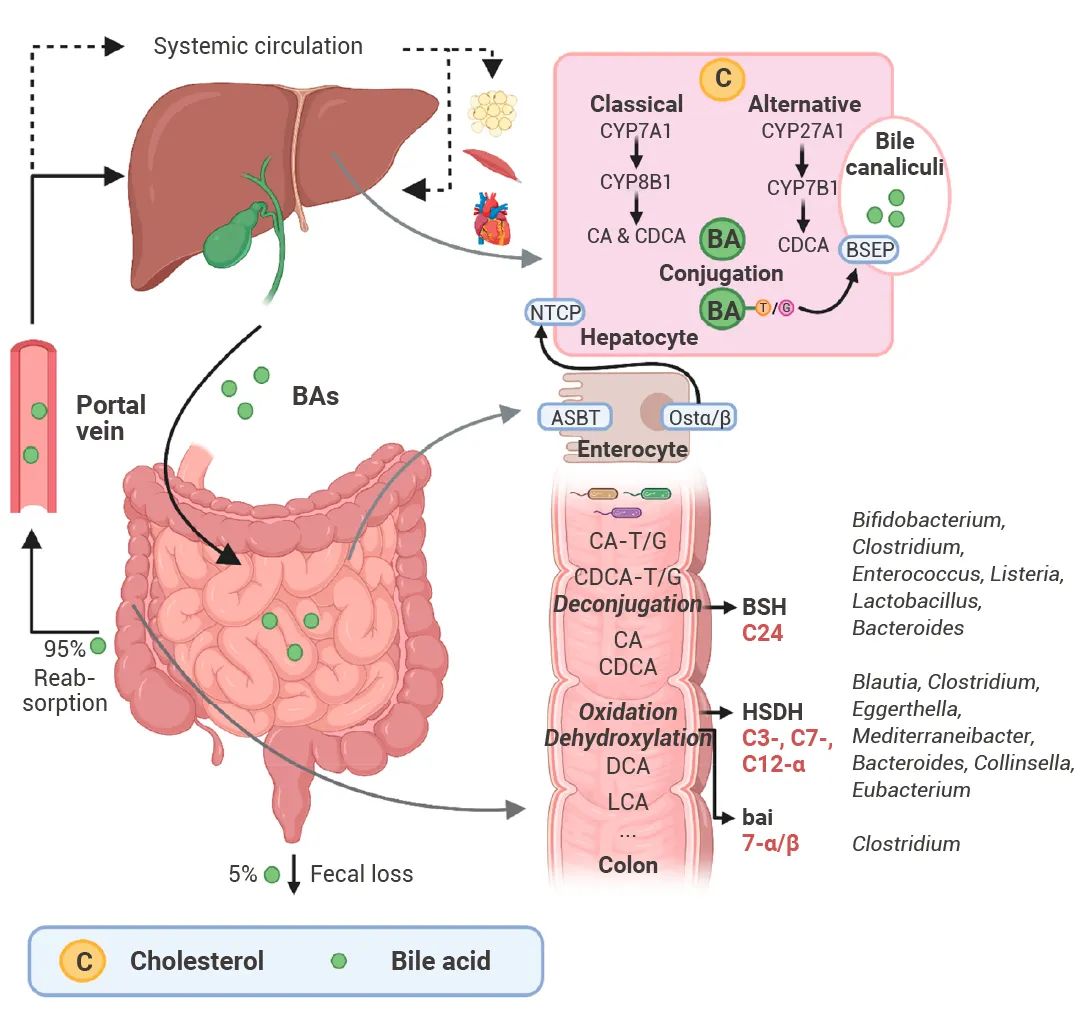
▐修饰宿主产生的代谢物进餐后,十二指肠刺激胆囊收缩,这时肝细胞中合成储存的初级 BAs 将排入肠道。在肠腔中,初级 BAs 可以溶解脂质,包括胆固醇和脂溶性维生素。95% 左右的初级 BAs 会经 ASBT 主动重吸收回到肝脏。 一小部分初级 BAs 逃逸并到达结肠,肠道微生物群可将初级 BAs 转化为次级 BAs,这一过程涉及三大类细菌酶,主要的结构修饰包括解共轭、羟基的差向异构化以及去结合和脱羟基等,从而导致 BAs 库多样化,并影响 BAs 信号传导[7]。
图 4. 肝脏 BA 合成、肠肝循环和体内微生物 BA 修饰[7]。 BA: bile acid; CA: cholic acid; CDCA: chenodeoxycholic acid; DCA: deoxycholic acid; UDCA: ursodeoxycholic acid; LCA : lithocholic acid; TCA: taurocholic acid; GCA: glycocholic acid; C: cholesterol; BSEP : bile salt-export pump; NTCP: Na+-taurocholic acid co-transporting polypeptide; ASBT : apical sodium-dependent BA transporter; Ostα/β: organic solute transporter α/β; BSH: bile salt hydrolases; HSDH : hydroxysteroid dehydrogenase; bai : BA inducible genes.
▐自身合成的代谢物 肠道菌群自身也可合成代谢物如 (1) 支链氨基酸 (Branched-chain Amino Acid, BCAA),可促进蛋白质合成并提供能量。(2) 多胺,能够与蛋白质、核酸类物质结合,调节细胞生长。(3) 维生素,在结肠被进一步利用[1]。这 3 类肠道菌群自身合成的物质,人体通过饮食也可获得。
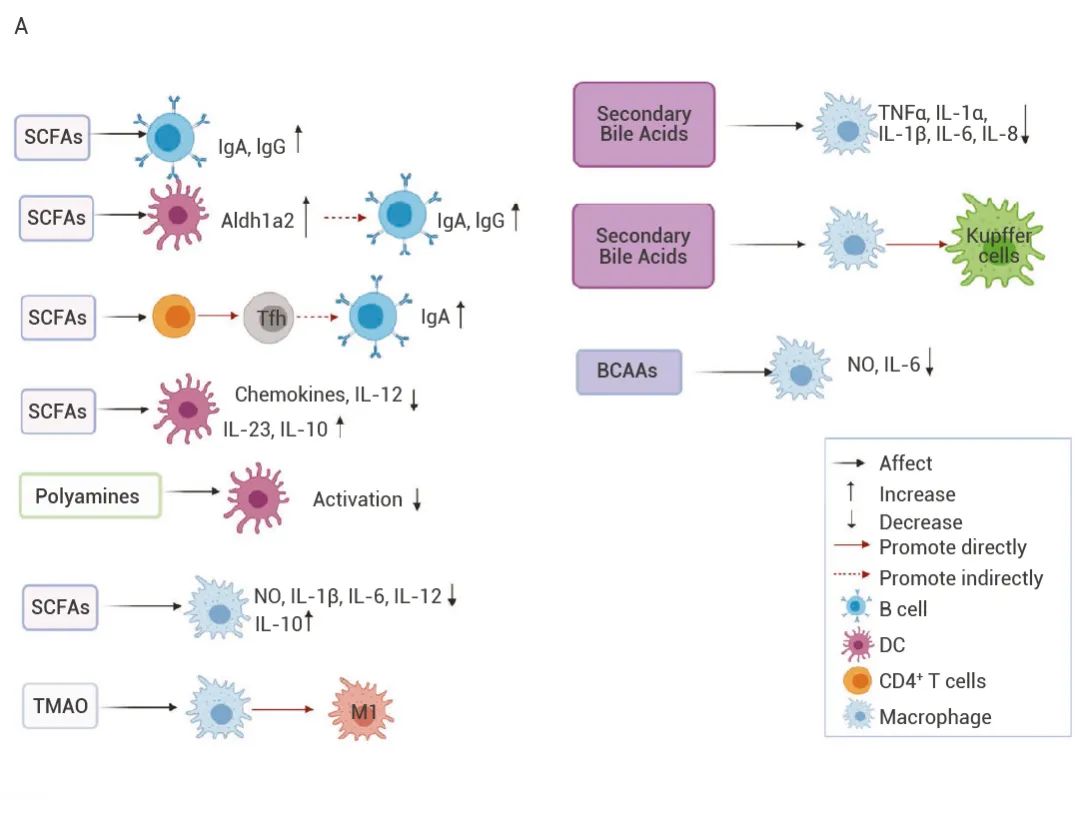
 ▐肠道菌群代谢物与肿瘤免疫 研究表明,70% 的免疫细胞生活在肠道中,肠道微生物群衍生的代谢物持续调节局部和全身的免疫细胞[1]。例如,增加肠道丁酸含量可促进 ILCs 和 CD4+ T 细胞产生 IL-22[8]。色氨酸代谢物 IE 等可显著抑制 NF-κB、IL-10R 表达[9]。以及石胆酸可降低 IL-1β、TNF-α、caspase-1 和 IL-22 的水平等[10]。 图 5. 肠道微生物群衍生的代谢物调节宿主免疫反应[1]。 (A) 肠道微生物群衍生的代谢物对 B 细胞、巨噬细胞和树突状细胞的影响; (B) 肠道微生物群衍生的代谢物对 T 细胞的影响。
而肿瘤免疫向来联系密切,一些微生物代谢物可通过塑造宿主免疫力来调节化疗和免疫治疗的抗肿瘤功效,进而影响癌症的发展。 清华大学医学院免疫所郭晓欢课题组发现肠道菌群通过其代谢产物短链脂肪酸丁酸,提高 CD8+ T 细胞中 ID2 的表达,增强 CD8+ T 细胞的抗肿瘤免疫应答,从而改善抗肿瘤治疗的效果[11]。
图 6. 肠道微生物代谢物通过调节细胞毒性 CD8+ T 细胞免疫促进抗癌治疗效果[11]。
Maik Luu 等人将 B16OVA 黑色素瘤细胞皮下注射到 CD45.2+ 小鼠体内,发现戊酸和丁酸通过代谢和表观遗传重编程增强细胞毒性 T 淋巴细胞 (Cytotoxic T-Lymphocyte, CTL) 的抗肿瘤活性[12]。此外,色氨酸代谢物—吲哚乙酸也可提高胰腺导管腺癌小鼠的化疗效果[13]。
本期小 M 为大家介绍了最近几年国自然研究较多的肠道菌群代谢物,其分类组成以及与肿瘤免疫的关联性。希望能让大家对肠道菌群代谢物有基础性的认识,有需要的小伙伴也可以点赞收藏喔~
 肠道菌群代谢化合物库
为了满足对肠道微生物的研究需求,MCE 精心挑选了 227 个肠道微生物代谢物。MCE 肠道微生物代谢物库将有助于肠道微生物研究及相关药物开发。
| Hyodeoxycholic acid
Hyodeoxycholic acid 是有肠道菌群在小肠中形成的次级胆汁酸,为 TGR5 (GPCR19) 的激动剂,在 CHO 细胞中,EC50 值为 31.6 µM。 | Xylose
D-(+)-木糖 (Xylose) 是一种天然化合物,经木糖异构酶催化形成木酮糖,这是木糖无氧乙醇发酵的关键步骤。 | Antrodin A
Antrodin A 是固态发酵樟脑菌菌丝体的主要活性成分之一。Antrodin A 通过提高肝脏的抗氧化和抗炎能力,维持肠道菌群的稳定性,保护肝脏免受酒精损伤。 | Dihydroferulic acid
Dihydroferulic acid (Hydroferulic acid) 是姜黄素的主要代谢产物之一,具有抗氧化/清除自由基的活性,IC50 值为 19.5 μM。Dihydroferulic acid 是人类肠道菌群的代谢产物,也是香草酸的前体物质。
| | |
[1] Yang W, et al. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol. 2021;18(4):866-877.
[2] Vinolo MA, et al. Regulation of inflammation by short chain fatty acids. Nutrients. 2011;3(10):858-76.
[3] Martin-Gallausiaux C, et al. SCFA: mechanisms and functional importance in the gut. Proc Nutr Soc. 2021;80(1):37-49.
[4] Roager HM, et al. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9(1):3294.
[5] Yang W, et al. Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseases. Cell Mol Immunol. 2021;18(4):866-877.
[6] Li X, et al. Trimethylamine-N-Oxide Pathway: A Potential Target for the Treatment of MAFLD. Front Mol Biosci. 2021;8:733507.
[7] Yntema T, et al. Emerging Roles of Gut Microbial Modulation of Bile Acid Composition in the Etiology of Cardiovascular Diseases. Nutrients. 2023;15(8):1850.
[8] Yang W, et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat Commun. 2020;11(1):4457.
[9] Scott SA, et al. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc Natl Acad Sci U S A. 2020;117(32):19376-19387.
[10] Shao J, et al. Synergistic anti-inflammatory effect of gut microbiota and lithocholic acid on liver fibrosis. Inflamm Res. 2022;71(10-11):1389-1401.
[11] He Y, et al. Gut microbial metabolites facilitate anticancer therapy efficacy by modulating cytotoxic CD8+ T cell immunity. Cell Metab. 2021 May 4;33(5):988-1000.e7.
[12] Luu M, et al. Microbial short-chain fatty acids modulate CD8+ T cell responses and improve adoptive immunotherapy for cancer. Nat Commun. 2021;12(1):4077.
[13] Tintelnot J, et al. Microbiota-derived 3-IAA influences chemotherapy efficacy in pancreatic cancer. Nature. 2023;615(7950):168-174.
| 




 楼主
楼主