登陆查看更多优秀资源帖,与同道便捷交流讨论
您需要 登录 才可以下载或查看,没有帐号?立即注册
x

提到 EMT,大多小伙伴们都会立刻想到肿瘤,我们常说,肿瘤具有 EMT 特性,但上皮-间充质转化 (EMT) 也是细胞形态发生过程中不可或缺的机制,因为没有间充质细胞,就永远无法形成组织和器官[1]。
EMT 过程描述了静止上皮细胞向间充质、运动表型的分化,最初在发育早期观察到[1]。后来人们发现 EMT 不仅对于发育和伤口愈合至关重要,而且代表了原发性肿瘤形成和转移的一个显著特性[2]。图 1. 上皮细胞可塑性的循环及诱导细胞 EMT 发生[1]。 A. 上皮细胞可以通过上皮-间充质转化过程转化为间充质细胞,在此期间上皮细胞失去了许多上皮特性,并具有间充质细胞的典型特性。在大多数生物发育的早期阶段,上皮间质转化 (EMT) 调控着重要的过程,在缺乏 EMT 的情况下,发育不能经过囊胚期。用 Matrix metalloproteinase-3 (C) 处理 SCp2 小鼠乳腺细胞 (B) 诱导 EMT。
EMT 过程可促进癌症干细胞样,免疫逃逸,多重抵抗,侵袭性表型,介导癌细胞的可塑性,允许其持续和不可逆地适应不断变化的条件,与肿瘤侵袭转移、耐药及免疫逃逸密不可分[3][4][5]。

EMT 受多种信号通路调节,例如 TGF-β、Wnt/β-catenin、Hedgehog 和 Notch 信号通路,这些途径通过刺激 Snail、Twist 和 ZEB1/2 等转录因子来触发 EMT。在所有信号通路中,Wnt/β-catenin 通路在 EMT 的调控中发挥着关键作用[6]。
▐Wnt/β-catenin 的激活诱发 EMT
正常情况下,Wnt/β-catenin 信号通路的负调控因子,如结肠腺瘤性息肉病蛋白(APC)、 糖原合成激酶3蛋白 (GSK-3) 和轴蛋白(Axin)等可与 β-catenin 结合形成复合物从而发生磷酸化,并进一步降解[7]。
图 2. 受抑制的 Wnt 信号级联[7]。
然而在多种癌症细胞内发现这些 Wnt/β-catenin 通路负调控因子的基因发生突变和缺失,β-catenin 在细胞质内大量聚集随后转移至核内与 T 细胞因子/淋巴增强因子 (Tcellfactor/Lymphoid enhancing factor, TCF/LEF) 等转录因子形成复合物,激活下游靶基因推动细胞周期发展或产生异常蛋白质,诱导 EMT,使细胞发生癌变[7]。
▐Wnt 通路的激活途径
Wnt 的激活是通过不同的 Wnt-蛋白质配体结合到 Frizzled 家族细胞表面受体,从而传递生物学信号至细胞内,包括三种途径[8]:
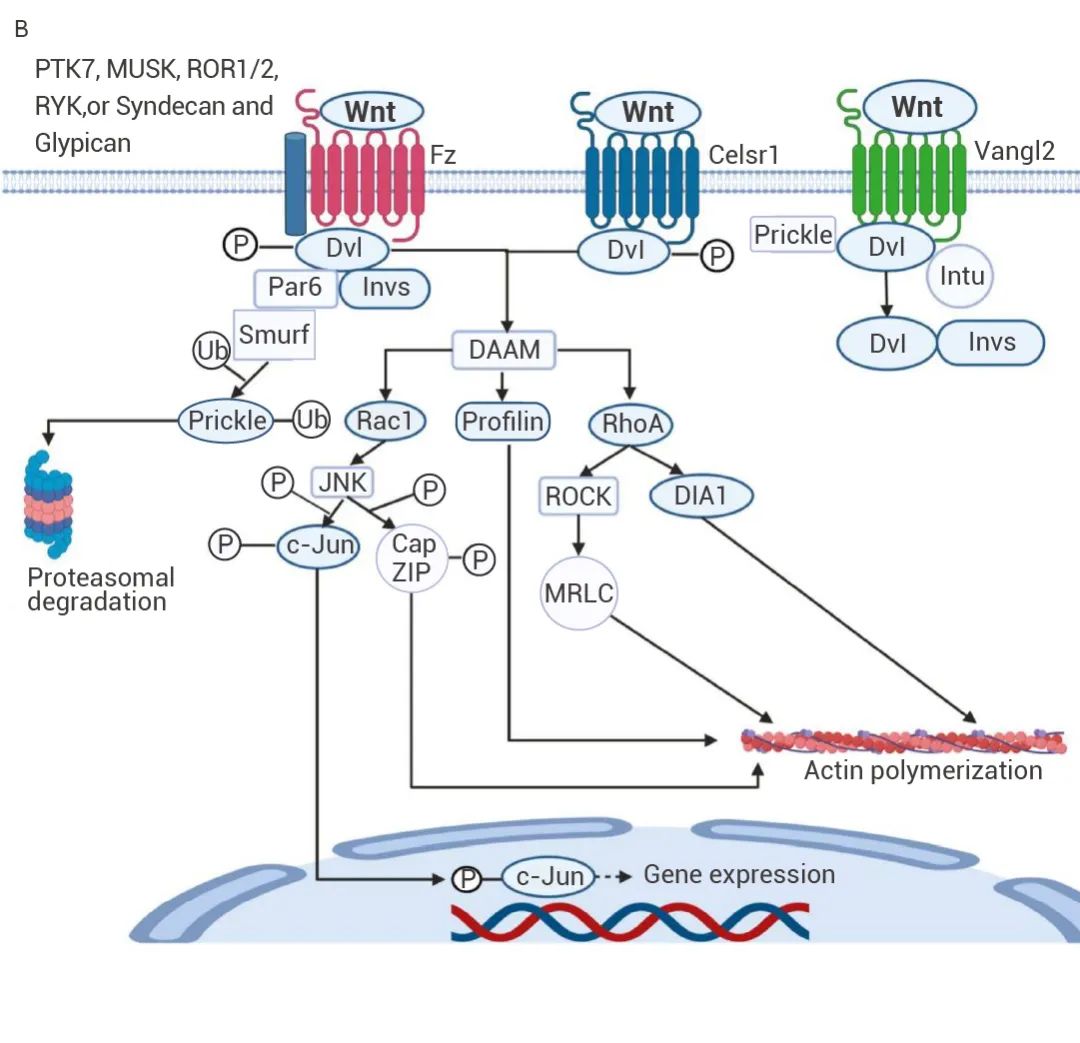
(1) 经典 Wnt/β-catenin 途径:Wnt 基因在肿瘤细胞中被异常激活,磷酸化激活胞质内的 Dsh 蛋白,抑制 GSK3β/APC/Axin 复合物中的关键成分 GSK3β 活性,阻止 GSK3β 对 β-catenin 的磷酸化与泛素化,降低了 β-catenin 的磷酸化降解[9]。胞质中聚集增多的 β-catenin 转进入细胞核,与核内转录因子 TCF/LEF 相互作用,从而影响细胞粘附、组织形态发生和肿瘤发展[10]。图 3. 经典 Wnt/β-catenin (A) 和非经典 Wnt/PCP 信号途径 (B)[7]。
(2) 非经典 Wnt/PCP 途径:一方面,Fz 受体结合 Wnt 配体并使 Dvl 磷酸化。另一方面,Smurf 泛素化 Prickle(一种通常抑制 Wnt/PCP 信号传导的蛋白质)。Prickle 的分解使 Dvl 能够与 DAAM 结合,激活 Rac1、Profilin 和 RhoA。Rac1 激活 JNK,从而磷酸化 c-Jun 和 CapZIP。然后 c-Jun 进入细胞核刺激基因转录。而 RhoA 激活 DIA1 和 ROCK,后者激活 MRLC。CapZIP、MRLC、DIA1 和 Profilin 都会刺激肌动蛋白聚合,进而影响细胞极性和迁移[7]。
(3) 非经典 Wnt/Ca2+ 途径:Wnt 与 Fz 受体结合导致 G 蛋白介导的 PLC 激活,刺激 Ca2+ 释放,DAG 与 Ca2+ 一起激活蛋白激酶 C (PKC) 以刺激细 Cdc 42,从而导致肌动蛋白聚合,从而促进细胞极化和迁移。同时,IP3 与 InsP3R 结合导致胞质 Ca2+ 增加,钙调神经磷酸酶激活活化 T 细胞的核因子 (NFAT),从而引起基因转录[7]。
图 4. 非经典 Wnt/Ca2+ 途径[7]。

大量研究表明,Wnt/β-catenin 信号通路的失调会导致 EMT,而 Wnt/β-catenin 的调控又有谁在参与呢?
▐miRNA 靶向 Wnt/β-catenin 来调节 EMT
Wnt/β-catenin 信号传导激活的核心是 β-catenin 在细胞质中的积累。因此,针对 β-catenin 的 miRNA 可能通过靶向 Wnt 信号通路或其下游转录因子来抑制 EMT。
今年 8 月,Dongsheng Zhu 等人揭示了miR-199b-3p在骨肉瘤 (OS) 的形成和进展中发挥关键作用。miR-199b-3p 可与 CCDC88A 的 3' 非翻译区 (UTR) 结合,下调 CCDC88A 的表达水平,抑制 EMT 和 Wnt/β-catenin 信号通路,从而介导其对 OS 细胞增殖和侵袭的抑瘤作用[11]。
图 5. miR-199b-3p/CCDC88A 轴在体外通过 Wnt/β-catenin 通路和 EMT 过程调节 OS 细胞的恶性行为[11]。
此外,Ling 等人的研究证明 miR-145 通过靶向 Oct4 使 Wnt/β-catenin 信号通路失活来抑制肺癌细胞中的 EMT[12]。miR-33b 还与 ZEB1 的 3′-UTR 结合并抑制肺腺癌细胞中的 ZEB1 表达,从而阻断 Wnt/β-catenin 信号传导并在体外和体内抑制肿瘤生长和 EMT 发生[13]。
▐WNT3A–RIP1–β-catenin 通路诱导 EMT
2023 年 7 月,A-Ram Kang 等人发现 了 RIP1 在控制 WNT/β-catenin 经典信号传导以增强结直肠癌 (CRC) 转移方面的新潜在作用[14]。
在没有 WNT 配体的情况下,磷酸化 β-catenin 被 β-TrCP 识别并结合。β-catenin 的泛素化、cIAP1/2 调节 RIP1 泛素化,进而降解 β-catenin 蛋白。
而 WNT3A 的处理诱导 cIAP1/2 降解,消除了 β-TrCP 向 β-catenin 的募集,并依次阻断 β-catenin 泛素化。RIP1 和 β-catenin 相互结合并稳定, RIP1 和 β-catenin 的这种结合也会刺激 β-catenin-β-TrCP 复合物的解离 (但不改变 β-catenin 和 β-TrCP 的蛋白水平),抑制 β-catenin 的泛素化,进而刺激 EMT 诱导,增强 CRC 细胞的体外迁移和侵袭能力[14]。
图 6. RIP1 在 CRC 转移中的作用模型[14]。
此外,许多致癌信号,如受体酪氨酸激酶 (RTKs) 家族、PI(3)K/Akt, MAPK 等可抑制 GSK-3β 活性,促使 β-catenin 的大量积累,触发细胞迁移和 EMT[15]。Wnt 家族会与 Ras/Raf/MEK/ERK,TGFβ/Smad 等通路产生 Crosstalk,相互影响协作调控与细胞侵袭转移相关的基因,共同参与调解肿瘤细胞 EMT 过程。
还有许多蛋白靶点通过参与调控 Wnt/β-catenin 而影响肿瘤细胞 EMT 过程。如 TAMs 分泌的白细胞介素-1β 通过磷酸化 GSK3β 在结肠癌细胞中增加 β-catenin 的可用性,阻碍 β-catenin 破坏复合体的功能[16]。MRGBP 通过 DKK1/Wnt/β-catenin 和 NF-κB/p65 通路介导 EMT 促进结直肠癌的进展等[17]。

本期小 M 为大家介绍了激活 Wnt/β-catenin 通路会诱发肿瘤 EMT 的产生,Wnt 通路的激活途径以及 Wnt/β-catenin 通路的相关调控。细胞的动态可塑性的调控也是研究者们寻求疾病治愈方法的重要突破点,涉及相关课题的小伙伴们可以点赞收藏喔~
 IWP-2
靶向结合 Porcupine ,Wnt 加工和分泌的抑制剂。 | XAV939
Tankyrase 1 抑制剂,靶向 Axin,抑制 Wnt 信号传导。 | IWR-1
IWR-1是端锚聚合酶抑制剂,抑制Wnt/β-catenin信号传导途径。 | SB-216763
ATP 竞争性的 GSK3 抑制剂。 | ICG-001
结合 CREB 蛋白的 β-catenin/TCF 转录抑制剂。 | MSAB
MSAB 是 Wnt/β-catenin 信号传导的有效和选择性抑制剂。MSAB 与 β-catenin 结合,促进其降解,并下调 Wnt/β-catenin 靶基因。 | NSC668036
Frizzled-Dvl 结构域抑制剂,阻断 Wnt 信号。 | Box5
Box5 是一种有效的 Wnt5a 拮抗剂。Box5 抑制 Wnt5a 信号并抑制 Wnt5a 介导的 Ca2+ 释放。Box5 抑制细胞迁移。 | TWS119
GSK-3β 抑制剂,激活 Wnt/β-catenin 通路。 | SKL2001
SKL2001 是 Wnt/β-catenin 信号通路的激动剂,具有抗肿瘤活性。SKL2001 通过破坏 Axin/β-catenin 相互作用,稳定细胞内 β-catenin。 |
 [1] Thiery JP, et al. Complex networks orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell Biol. 2006 Feb, 7(2):131-42. [2] Bakir B, et al. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020 Oct, 30(10):764-776. [3] Yeung KT, et al. Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol. 2017 Jan, 11(1): 28-39. [4] Brabletz S, et al. Dynamic EMT: a multi-tool for tumor progression. EMBO J. 2021 Sep 15, 40(18): e108647. [5] Hay ED. An overview of epithelio-mesenchymal transformation. Acta Anat (Basel). 1995, 154(1): 8-20. [6] Lei Y, et al. MicroRNAs target the Wnt/β‑catenin signaling pathway to regulate epithelial‑mesenchymal transition in cancer (Review). Oncol Rep. 2020 Oct, 44(4): 1299-1313. [7] Qin K, et al. Canonical and noncanonical Wnt signaling: Multilayered mediators, signaling mechanisms and major signaling crosstalk. Genes Dis. 2023 Mar 24, 11(1): 103-134. [8] Buyuk B, et al. Epithelial-to-Mesenchymal Transition Signaling Pathways Responsible for Breast Cancer Metastasis. Cell Mol Bioeng. 2021 Sep 2, 15(1): 1-13. [9] Yu F, et al. Wnt/β-catenin signaling in cancers and targeted therapies. Signal Transduct Target Ther. 2021 Aug 30, 6(1): 307. [10] Lecarpentier Y, et al. Multiple Targets of the Canonical WNT/β-Catenin Signaling in Cancers. Front Oncol. 2019 Nov 18, 9: 1248. [11] Zhu D, et al. hsa-miR-199b-3p suppresses osteosarcoma progression by targeting CCDC88A, inhibiting epithelial-to-mesenchymal transition, and Wnt/beta-catenin signaling pathway. Sci Rep. 2023 Aug 2, 13(1): 12544. [12] Ling DJ, et al. MicroRNA-145 inhibits lung cancer cell metastasis. Mol Med Rep. 2015 Apr, 11(4): 3108-14. [13] Qu J, et al. MicroRNA-33b inhibits lung adenocarcinoma cell growth, invasion, and epithelial-mesenchymal transition by suppressing Wnt/β-catenin/ZEB1 signaling. Int J Oncol. 2015 Dec, 47(6): 2141-52. [14] Kang AR, et al. A novel RIP1-mediated canonical WNT signaling pathway that promotes colorectal cancer metastasis via β -catenin stabilization-induced EMT. Cancer Gene Ther. 2023 Jul 27. [15] Zhou, et al. Dual regulation of Snail by GSK-3β-mediated phosphorylation in control of epithelial–mesenchymal transition. Nat Cell Biol. 2004, 6: 931-940. [16] Kaler P, et al. Macrophage-derived IL-1beta stimulates Wnt signaling and growth of colon cancer cells: a crosstalk interrupted by vitamin D3. Oncogene. 2009 Nov 5, 28(44): 3892-3902. [17] Long X, et al. MRGBP promotes colorectal cancer metastasis via DKK1/Wnt/β-catenin and NF-kB/p65 pathways mediated EMT. Exp Cell Res. 2022 Dec 1, 421(1): 113375.
| 




 楼主
楼主