登陆查看更多优秀资源帖,与同道便捷交流讨论
您需要 登录 才可以下载或查看,没有帐号?立即注册
x
本帖最后由 BLZJ_7d91db 于 2024-2-21 11:08 编辑
 Pyroptosis,即细胞焦亡,又称炎症程序性细胞死亡。小伙伴们可能要说了,“Wow~字都认识,话是真没听懂”。 首先呢,它是一种细胞程序性死亡 (Programmed Cell Death, PCD),即细胞接受某种信号或受到某些因素刺激后,为了维持内环境稳定,按照预定的程序发生的一种主动性消亡过程。也就是说,这是完全自愿的高尚牺牲!
程序性细胞死亡主要包括:程序性坏死(Necroptosis)、凋亡(Apoptosis)、细胞焦亡(Pyroptosis)和铁死亡(Ferroptosis)等。感兴趣的小伙伴可以参考小 M 往期的细胞死亡相关推文,此处就不再赘叙。
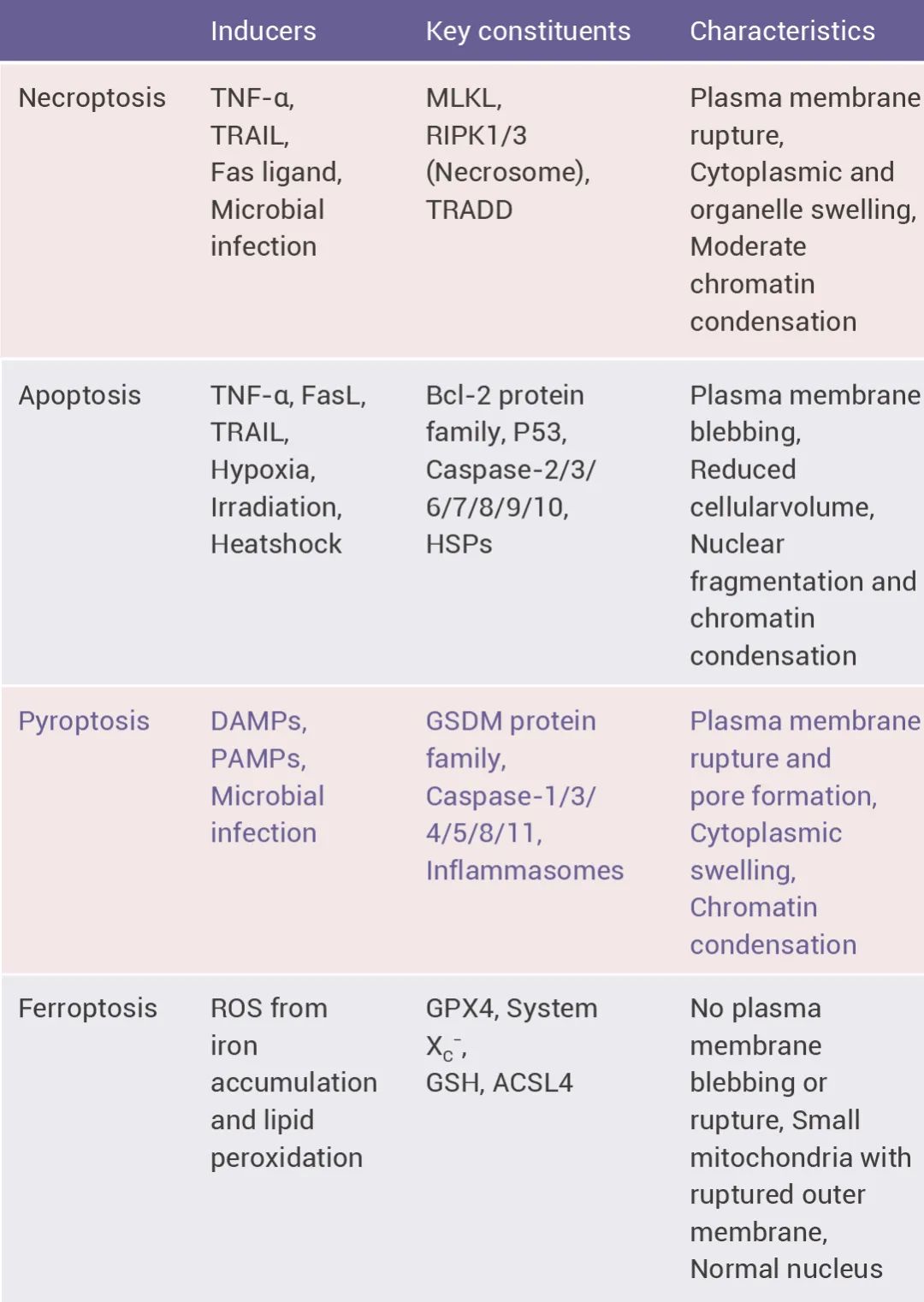
其次,细胞焦亡是一种炎症形式,其由促炎信号触发并与炎症相关[1],可经微生物感染或宿主因素引发,最常在细胞内细菌或病原体感染时发生[2]。它是由炎症小体的活化及炎症性半胱天冬酶 (Caspases) 的激活引起的(炎症小体往期回顾:炎症小体:免疫炎症领域热门选手)。细胞焦亡会发生细胞肿胀、细胞膜起泡等细胞溶解现象(类似坏死),涉及多种以炎症和细胞死亡为特征的疾病,包括自身炎症综合征、神经退行性疾病、炎症性肠病和脑缺血等。 图 1. 细胞焦亡与其他细胞程序性死亡的特征[3]。
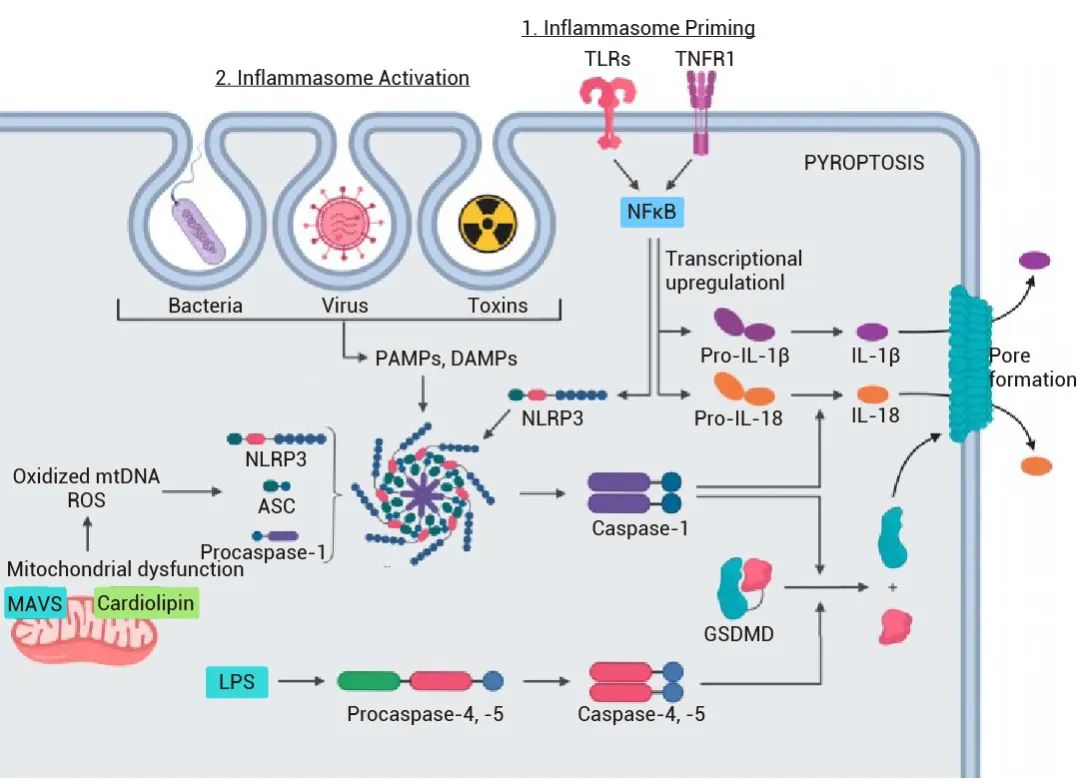
▐ 细胞焦亡过程[4]:细胞焦亡与细胞凋亡和细胞坏死不同,它是由炎症小体的活化及炎症性半胱天冬酶的激活引起的,通常与感染、疾病和免疫反应有关。细胞焦亡的过程涉及多个环节,如病原体感染、炎性小体的形成与活化等[4]。
图 2. 细胞焦亡概述[5]。
 如下所述 如下所述
1. 病原体感染:病原体进入宿主细胞,被宿主细胞的模式识别受体 (PRRs) 识别,如 Toll 样受体 (TLRs)、NOD 样受体 (NLRs) 和 RIG-I 样受体 (RLRs) 等。
2. 炎症小体形成:识别到病原体的细胞会激活炎性小体。典型的炎性小体包括 NLRP3、ASC 和 Caspase-1。激活后的 NLRP3 与 ASC 结合,继而招募 Caspase-1,形成炎性小体。
3. Caspase 蛋白活化:炎性小体的形成将导致 Caspase-1 的活化。活化的 Caspase-1 能够切割并激活一系列底物,如炎症性细胞因子前体 (如 pro-IL-1β 和 pro-IL-18) 和 Gasdermin D。
4. Gasdermin D 活化:活化的 Caspase-1 切割 Gasdermin D,产生 N 端和 C 端两个片段。其中活性的 N 末端片段可以穿透细胞膜,形成孔道。细胞膜孔道的形成导致细胞内液体和离子的快速流动,使细胞发生肿胀。
5. 细胞膜破裂与炎症:膜孔道的形成会致使细胞破裂。在这个过程中,细胞被迫吸收外界离子和水分,同时向外释放细胞内容物,如炎症因子 IL-1β 和 IL-18。这些炎症因子将进一步引发免疫系统的炎症反应。
6. 细胞碎片及清理:聚集在焦亡细胞附近的周围的巨噬细胞会清理最后的战场,清除细胞碎片,帮助减轻炎症反应。
 细胞焦亡主要依靠炎症小体激活 Caspase 家族蛋白引起各种生理反应[6]。而炎症小体通常由传感器蛋白、衔接蛋白 (ASC)、以及无活性的 Caspase 蛋白前体组成。这种复合物触发的焦亡可分为 Caspase-1 激活的经典途径或 Caspase-11/4/5 激活的非经典途径[7]。 ▐ 经典焦亡:Caspsae-1[4][8][9]: Caspase-1 是焦亡过程中最重要的 Caspases 家族成员,通过集结响应不同刺激的炎性传感因子,构建出了专属 Caspase-1 经典通路战舰群:NLRP3 炎性体、AIM2 炎性体、NLRP1 炎性体、PYRIN 炎性体,以及 NLRC4 炎性体。这些炎症小体可介导底物 Gasdermin D (GSDMD) 或炎症因子前体 (如 pro-IL-1β 和 pro-IL-18) 的裂解,打通膜孔道,而这类变化将导致细胞膜破裂,细胞释放炎症因子,引发炎症反应,激发焦亡[8]。
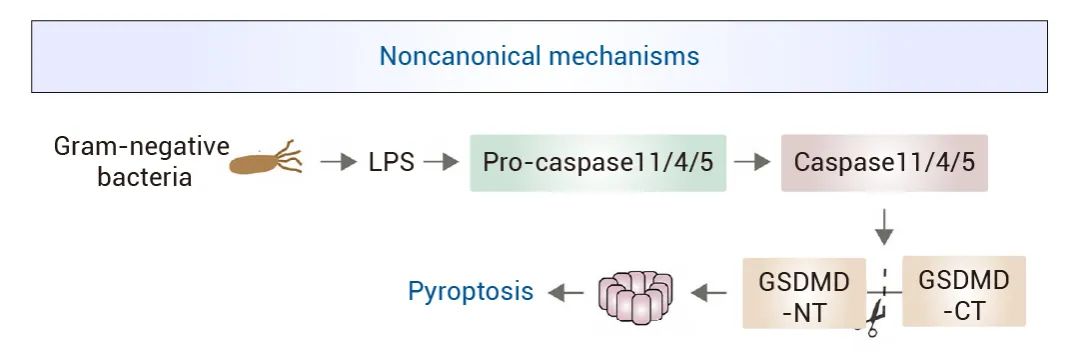
图 3. Caspase-1 激活的经典途径[9]。 ▐ 非经典焦亡:Caspase-11/4/5[6][10]: 比起规模庞大,训练有素的 Caspase-1 战队,Caspase-11/4/5 更像伺机而动的侦察兵。它们可以直接识别病原体的脂多糖 (LPS),从而通过非炎性小体途径激活焦亡过程。当 Caspase-11/4/5 与 LPS 结合时,可直接切割 GSDMD,产生活性 N 末端片段,形成细胞膜孔道,导致焦亡。 图 4. Caspase-11/4/5 激活的非经典途径[9]。
在一些关键时刻,Caspase-4/11 在肺动脉高压 (PAH) 动物模型和 TNF-α 诱导的人肺动脉内皮细胞中被激活。活化的 Caspase-4/11 还能够剪切 Caspase-3 前体,使其活化。被激活的 Caspase-3 在随后裂解 Gasdermin E (GSDME),造成细胞膜穿孔,进一步推动焦亡。而 Caspase-5 在某些情况下可以参与炎性小体的组装和激活,发挥类似类似 Caspase-1 的作用进而影响焦亡途径。 ▐ Caspase-3 引发的细胞焦亡[11]: 在上一个单元故事中早早露面的 Caspase-3 并不神秘。它主要与细胞凋亡过程有关,而近年来,人们发现了 Caspase-3 的另一个作用,它可以位于 GSDME 的上游,在焦亡中发挥炎症切割作用[12]。 图 5. caspase-3 细胞凋亡以及 caspase-3/GSDME 介导的细胞焦亡途径[12]。
在病毒感染的背景下,Caspase-3 被激活并间接促使 GSDME 的切割。GSDME 被剪切为 N 端和 C 端两个片段,其中 GSDME-N 端片段在细胞膜上形成孔道,导致离子和水分进入细胞,使细胞膜破裂。
在一些情况下,活化的 Caspase-1 导致细胞内线粒体损伤后,也会诱发 Caspase-3 的激活,推动焦亡。 ▐ Caspase-6/8/9与细胞焦亡: Caspase-6、Caspase-8 和 Caspase-9 的不同调节也会影响焦亡途径。小 M 为大家整理在下面啦~ 图 6. Caspase-6/8/9 对细胞焦亡的影响。 
细胞焦亡如何检测呢?下面小 M 以文献为例为大家介绍几种常用的细胞焦亡检测方法~
近期发表的文献 "Mannose antagonizes GSDME-mediated pyroptosis through AMPK activated by metabolite GlcNAc-6P" 表明,CCCP (Carbonyl cyanide m-chlorophenyl hydrazine) 加铁可诱导黑色素瘤细胞中 GSDME 依赖性细胞焦亡,而甘露糖可逆转不同癌细胞系中 GSDME 介导的细胞焦亡[17]。
作者使用甘露糖预处理不同的癌细胞系,然后用不同的诱导剂处理 24 小时,检测评估了焦亡的发生:包括特征形态、GSDME 裂解和 LDH 释放。
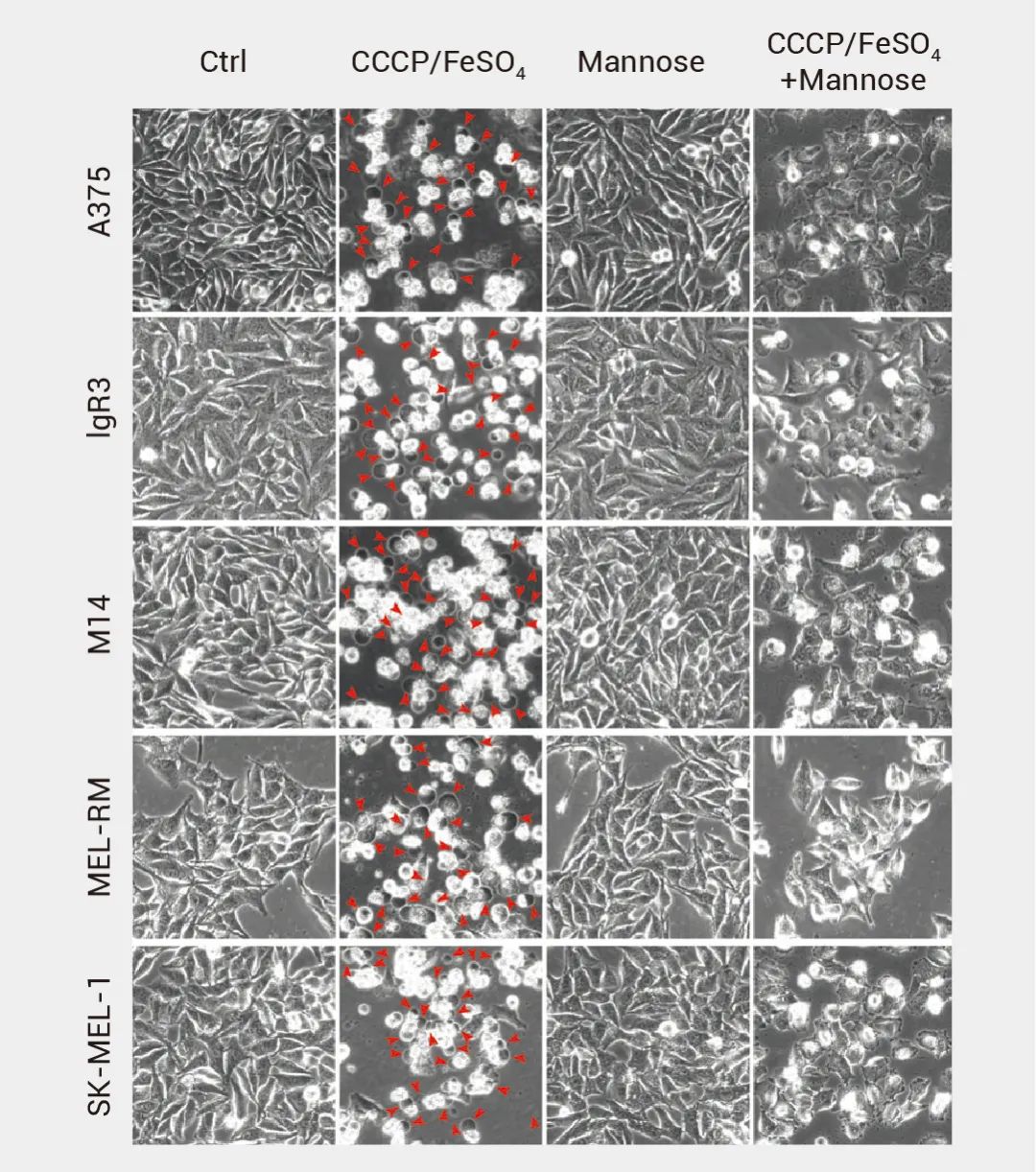
▐ 形态观察:
实时监测细胞状态和细胞死亡,分辨焦亡引起的细胞形态变化,如细胞质肿胀和膜破裂等。图 7. 焦亡细胞的形态变化 (红色箭头表示焦亡细胞)[17]。 ▐ Caspase、Gasdermins 蛋白的裂解[17]: 细胞焦亡期间会发生 Gasdermins 和Caspases 蛋白的剪切,可通过免疫印迹 (Western blot) 等确定蛋白的裂解。例如,使用甘露糖预处理黑色素瘤细胞系,后用 CCCP/FeSO4 诱导焦亡,可观察到焦亡细胞中发生 GSDME 裂解[17]。 图 8. 焦亡细胞中发生 GSDME 裂解[17]。
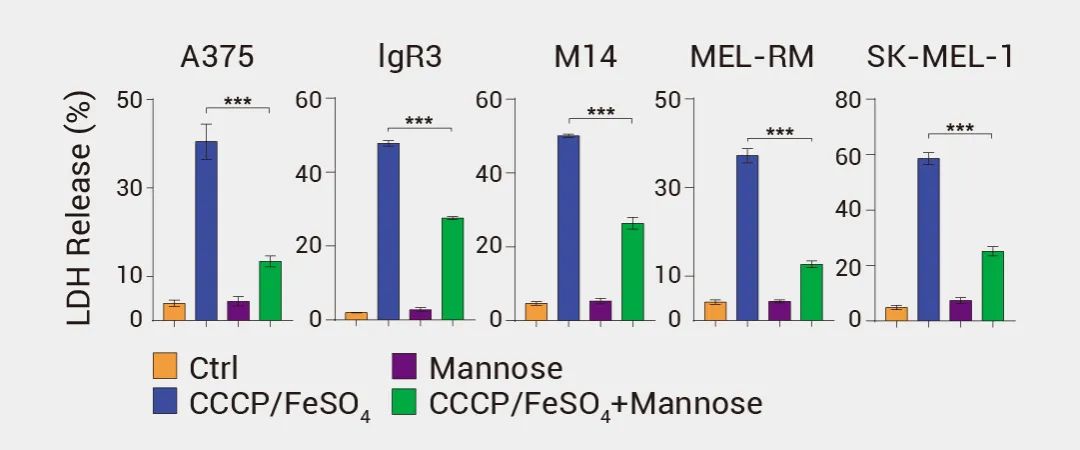
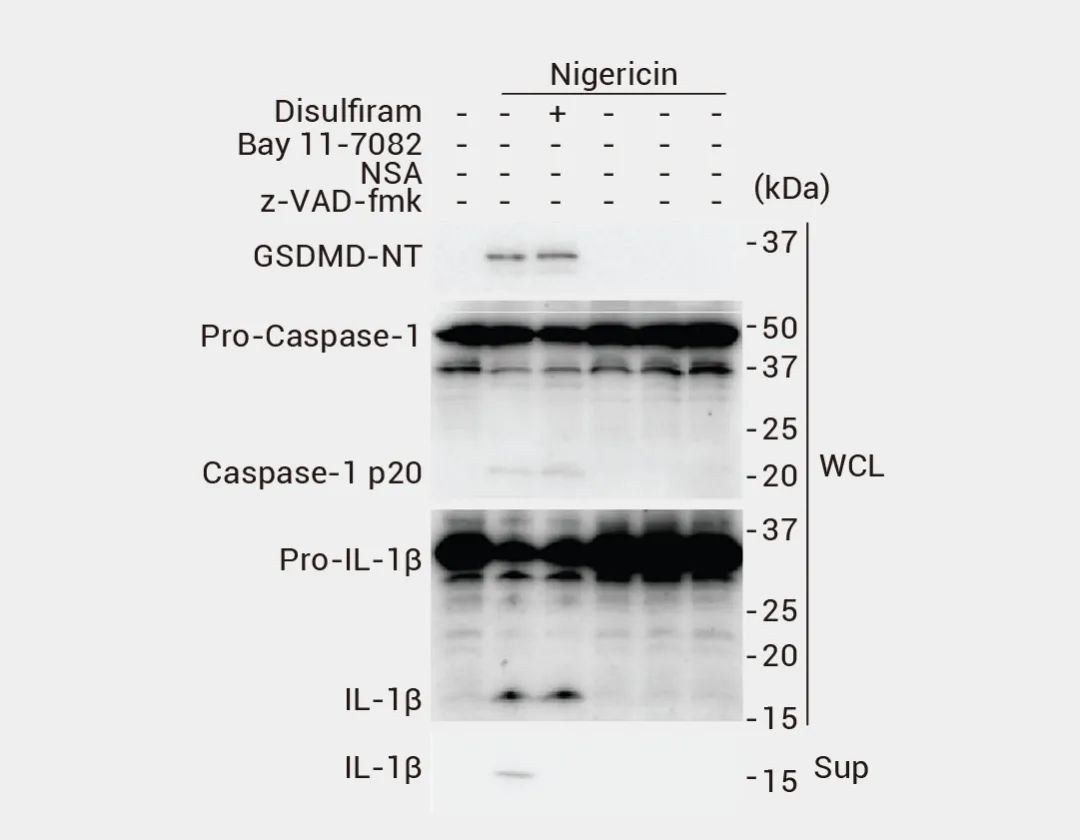
▐ LDH 释放实验:当细胞发生焦裂时,细胞膜会破裂并释放 LDH。可以通过检测培养液中 LDH 活性的变化来判断细胞是否发生焦裂[16]。作者使用甘露糖预处理不同的黑色素瘤细胞系,后用 CCCP/FeSO4 诱导焦亡,结果表明,与对照组(Ctrl)相比,CCCP/FeSO4 实验组 LDH 大量释放[17]。 图 9. 焦亡细胞中乳酸脱氢酶 (LDH) 的释放[17]。 ▐ IL-1β 和 IL-18 释放: 炎性程序性细胞死亡通常伴随着炎症因子 IL-1β 和 IL-18 的释放。可通过ELISA 或其他免疫检测方法检测这些炎症因子的释放情况[17]。例如,LPS 引发的 THP-1 细胞在没有抑制剂的情况下用 Nigericin 处理时,检测到 Caspase-1、GSDMD 和 pro-IL-1β 裂解为对应的活性形式。图 10. pro-IL-1β 的裂解和 IL-1 的释放[17]。 细胞焦亡通常伴随着孔素的形成和细胞膜的破裂。可以使用荧光染料 (如丙啶酮、YO-PRO-1 或 PI) 标记孔素并通过流式细胞术或荧光显微镜观察细胞膜孔的形成[17][18]。
此外,还可通过炎性小体形成和炎性半胱天冬酶的活化来判断:细胞焦亡的特点是炎性体的形成,其标记物是 NLRP3。因此,可通过免疫印迹(Western blot)、流式等技术检测炎性小体 NLRP3 的表达和 Caspase-1 的活化情况[19]。

Disulfiram
Disulfiram (双硫仑) 是特异性的乙醛脱氢酶 1 型 (ALDH1) 的抑制剂,可抑制细胞焦亡 (pyroptosis)。Disulfiram 可抑制人体和小鼠细胞中脂质体中的 GSDMD 孔形成和 IL-1β 分泌。 | LDC7559
LDC7559 是 GSDMD 的抑制剂,抑制中性粒细胞胞外杀菌网络 (NET) 后期阶段。LDC7559 可直接阻断 GSDMD NT 域的致死活动,抑制细胞焦亡 (pyroptosis)。 | Morroniside
Morroniside (莫诺苷) 是 MMP2/9 的抑制剂,并显著降低活性 caspase-3水平。Morroniside 通过调节 Bax/Bcl-2 的比例增加而显示出对细胞焦亡 (pyroptosis) 的抑制活性。 | Ac-FLTD-CMK
Ac-FLTD-CMK 是 GSDMD 衍生的抑制剂,特异性抑制炎症性 caspases 1/4/5/11,IC50=46.7 nM (caspases-1),1.49 nM (caspases-4),329 nM (caspases-5)。Ac-FLTD-CMK 抑制 GSDMD 的切割过程,不靶向 caspase-3 或细胞凋亡。 | ICy OH
ICy-OH,是一种化学抗癌剂和碘化光敏剂 (λex=640 nm,λem=690-740 nm)。ICy-OH 还可以诱导胰腺癌细胞发生焦亡 (pyroptosis)。 | Cleaved-Caspase 3 p12 Antibody
Cleaved-Caspase 3 p12 Antibody 是一个非偶联、兔源、抗 Cleaved-Caspase 3 p12 单克隆抗体。它可用于人、小鼠、大鼠背景下 WB、ICC/IF 实验,且不带标记。 | beta Tubulin Antibody
beta Tubulin Antibody 是一个非偶联、分子量约 50 kDa、兔源、抗 beta Tubulin 单克隆抗体。它可用于人、小鼠、大鼠背景下 WB、ICC、IHC-P、FC 实验,且不带标记。 |
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
【1】SW Ryter, et al. Cell Death and Repair in Lung Disease. Pathobiology of Human Disease. 2014, Pages 2558-2574.
【2】Miller DR, et al. The interplay of autophagy and non-apoptotic cell death pathways. Int Rev Cell Mol Biol. 2020;352:159-187.
【3】Loveless R,et al. Pyroptosis at the forefront of anticancer immunity. J Exp Clin Cancer Res. 2021 Aug 24;40(1):264.
【4】Si Ming Man, et al. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases.Immunol Rev. 2017 May;277(1):61-75.
【5】Del Re DP, et al. Fundamental Mechanisms of Regulated Cell Death and Implications for Heart Disease. Physiol Rev. 2019 Oct 1;99(4):1765-1817.
【6】Yu P, et al. Pyroptosis: mechanisms and diseases. Signal Transduct Target Ther. 2021 Mar 29;6(1):128.
【7】Van Opdenbosch N, et al. Caspases in Cell Death, Inflammation, and Disease. Immunity. 2019 Jun 18;50(6):1352-1364.
【8】Rathinam V A K, et al. Regulation of inflammasome signaling[J]. Nature immunology, 2012, 13(4): 333-342.
【9】Wei X, et al. Role of pyroptosis in inflammation and cancer. Cell Mol Immunol. 2022 Sep;19(9):971-992.
【10】Wu Y, et al. Caspase-4/11-Mediated Pulmonary Artery Endothelial Cell Pyroptosis Contributes to Pulmonary Arterial Hypertension. Hypertension. 2022 Mar;79(3):536-548.
【11】Wang Y, et al. Chemotherapy drugs induce pyroptosis through Caspase-3 cleavage of a gasdermin. Nature. 2017 Jul 6;547(7661):99-103.
【12】Jiang M, et al. The caspase-3/GSDME signal pathway as a switch between apoptosis and pyroptosis in cancer. Cell Death Discov. 2020 Oct 28;6:112.
【13】Sheng M, et al. Caspase-6/NR4A1/SOX9 signaling axis regulates hepatic inflammation and pyroptosis in ischemia-stressed fatty liver. Cell Death Discov. 2023 Mar 28;9(1):106.
【14】Fritsch M, et al. Caspase-8 is the molecular switch for apoptosis, necroptosis and pyroptosis. Nature. 2019 Nov;575(7784):683-687
【15】Zheng Z, et al. The Lysosomal Rag-Ragulator Complex Licenses RIPK1 and Caspase-8-mediated Pyroptosis by Yersinia. Science. 2021 Jun 25;372(6549):eabg0269.
【16】Li T, et al. Aloe-Emodin Induces Mitochondrial Dysfunction and Pyroptosis by Activation of the Caspase-9/3/Gasdermin E Axis in HeLa Cells. Front Pharmacol. 2022 May 17;13:854526.
【17】Ai YL, et al. Mannose antagonizes GSDME-mediated pyroptosis through AMPK activated by metabolite GlcNAc-6P. Cell Res. 2023 Jul 17.
【18】Hu JJ, et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nat Immunol. 2020 Jul;21(7):736-745.、
【19】Kucia M, et al. An evidence that SARS-Cov-2/COVID-19 spike protein (SP) damages hematopoietic stem/progenitor cells in the mechanism of pyroptosis in Nlrp3 inflammasome-dependent manner. Leukemia. 2021 Oct;35(10):3026-3029.
| 




 楼主
楼主